Gliia (vanakreeka sõnast γλία, γλοία 'liim') ehk neurogliia on närvisüsteemi kude, millel on perifeerses ja kesknärvisüsteemis mitu tugifunktsiooni, sealhulgas närvijätkete elektriline isoleerimine, rakuvälise keskkonna reguleerimine ja kaitsefunktsioon.
Gliiarakkudeks ehk gliotsüütideks nimetatakse närvisüsteemi rakke, mis ei ole närvirakud. Gliiarakke on mitut tüüpi. Need erinevad omavahel funktsiooni, ehituse ja asukoha poolest.
Esimesena kirjeldas gliia rakke saksa patoloog Rudolf Virchow 1856. aastal.
Gliiarakud moodustavad kesknärvisüsteemi ruumalast poole. Inimese ajus on umbes 86 miljardit närvirakku ja ligikaudu 85 miljardit gliiarakku, kusjuures gliiarakkude ja närvirakkude vahekord on aju eri osades oluliselt erinev. Näiteks ajukoores on gliiarakke umbes 3,7 korda rohkem kui närvirakke, samas kui väikeajus on närvirakke gliiarakkudest üle nelja korra rohkem.
Funktsioon
Gliial on teada viis põhilist funktsiooni:
- Struktuurne funktsioon.
- Närvirakkude varustamine toitainete ja hapnikuga.
- Rakuvälise keskkonna reguleerimine – gliiarakud reguleerivad rakuvälist ioonide kontsentratsiooni ning eritavad ja eemaldavad neurotransmittereid.
- Närvirakkude ja närvijätkete isoleerimine – gliiarakud moodustavad närvijätkete ümber müeliintupe, mis võimaldab närvisignaalide kiiremat kulgemist.
- Haigustekitajate hävitamine ja surnud närvirakkude eemaldamine – teatud gliiarakud on võimelised fagotsütoosiks.
Gliiarakud sünteesivad neurosteroide.
Gliiarakkude tüübid
Mikrogliia
 Mikrogliiarakud pärinevad
Mikrogliiarakud pärinevad - mesodermist ning täidavad kesknärvisüsteemis makrofaagide
- funktsiooni. Erinevalt närvirakkudest liiguvad mikrogliiarakud ringi. Need jälgivad aju sisekeskkonda ning sooritavad vajaduse korral fagotsütoosi.
Mikrogliia rakud ekspresseerivad D1- ja D2-dopamiiniretseptoreid. Dopamiin, toimides D1-le, reguleerib mikrogliia lämmastikoksiidi sünteesi.
Ajutrauma või infektsiooni korral mikrogliia rakud jagunevad ning vahendavad vajalikke kaitsefunktsioone.

Mikrogliia roti ajukoores (lektiini värving)

Makrogliia
- Makrogliiarakud arenevad ektodermist ehk välisest lootelehest.
- Astrotsüüdid (vahel ka astrogliia) on arvukaim gliiarakkude tüüp kesknärvisüsteemis. Astrotsüüdid on nimetuse saanud selle järgi, et nad on "tähekujulised". Astrotsüüdid reguleerivad närvisüsteemi keemilist keskkonda, kohandades ioonide kontsentratsiooni ning eemaldades sünaptilisel ülekandel eralduvaid neurotransmittereid.
- Oligodendrotsüüdid isoleerivad kesknärvisüsteemis närvirakkude aksoneid, moodustades müeliintupe, mis võimaldab kiiremat närvisignaalide kulgu.
- Ependüümirakud (vahel ka ependümotsüüdid) ümbritsevad seljaaju ja ajuvatsakeste sisemust. Ependüümirakud sünteesivad aju-seljaajuvedelikku ning kontrollivad ainevahetust liikvori ja närvisüsteemi vahel. Ependüümirakud võivad teatud tingimustes käituda närvikoe tüvirakkudena.
- Radiaalgliia mängib olulist rolli närvisüsteemi arengus, kus radiaalgliiarakud moodustavad juhteteed, mida mööda migreeruvad diferentseeruvad närvirakud. Radiaalgliiarakud käituvad ka kui närvirakkude eellasrakud, diferentseerudes hilisemas arengus närvirakkudeks. Täiskasvanu ajus võib radiaalgliiarakke täheldada väikeajus (ehk tserebellumis) ja silma võrkkestas. Tserebellumis nimetatakse radiaalgliiarakke Bergmanni gliiaks, mis reguleerib sünaptilist plastilisust. Silma võrkkestas on radiaalsed Mülleri gliiarakud peamiseks gliiarakutüübiks, mis täidavad gliiale omaseid tugifunktsioone.
- Schwanni rakud täidavad oligodendrotsüütidega sarnast rolli perifeerses närvisüsteemis. Schwanni rakud moodustavad perifeersete närviaksonite ümber müeliintupe, mis võimaldab kiiremat närvisignaalide levikut. Perifeerse närvi vigastamisel võivad Schwanni rakud muutuda fagotsütootiliseks ning toota mitmeid perifeerse närvi regenereerumiseks vajalikke signaale.
Gliiarakkude vahekorrad erinevad oluliselt sõltuvalt indiviidist. Nii näiteks tuvastati ühes uuringus eri vanuses inimeste neokorteksirakkude steroloogilise loendamise käigus, et gliiarakkudest on keskmiselt 75,6% oligodendrotsüüdid, 17,3% astrotsüüdid ja 6,5% mikrogliia ning vanuse kasvades kahaneb oligodendrotsüütide arv. Samuti leiti, et meestel on gliiarakude arv suurem kui naistel.
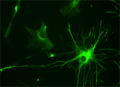
Inimese astrotsüüt rakukultuuris

Oligodendrotsüüt, mis moodustab mitme aksoni ümber müeliintupe






