Intron on geenis olev nukleotiidne järjestus, mis eemaldatakse RNA splaissimise käigus, kui valmistatakse RNA produkt, millelt toimub translatsioon. Intronid ja eksonid geenis transkribeeritakse RNA molekuliks ning eksonid liidetakse kokku RNA lõpp-produktiks, kui intronid on eemaldatud. Intron on geeni mittekodeeriv piirkond.
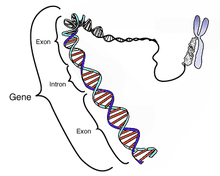
Üht intronit sisaldav geen
Terminit intron kasutatakse nii DNA sees olevate intronite iseloomustamiseks kui ka RNA sees olevate intronite iseloomustamiseks.
Järjestused, mis hiljem kokku liidetakse RNA valmistamiseks, on moodustunud eksonitest pärast RNA splaissimist. Intronid esinevad enamiku organismide geenides ning ka paljudes viirustes. Introneid esineb paljudes geenides, ka neis, mis genereerivad valke, ribosomaalset-RNAd (rRNA) ja transport-RNAd (tRNA). RNA töötlemine toimub enne translatsiooni ning pärast transkriptsiooni.
Sõna intron tuleb ingliskeelsest väljendist intragenic region (geenisisene piirkond), mis tähendab geeni sees olevat ala.
KlassifikatsioonOn teada vähemalt nelja eri tüüpi introneid.
Pakutakse ka grupp III introneid omaette perekonnana, kuid neist teatakse veel vähe.
Grupp I ja grupp II introneid leidub valke kodeerivate geenides (informatsiooni-RNA ehk mRNA), transport-RNA ja ribosomaalses RNAs paljudel organismidel. Pärast RNA transkriptsiooni teevad I grupi ja II grupi intronid läbi ulatusliku sisemise interaktsiooni, mis lubab neil kolmedimensiooniliseks kujundiks kokku voltuda. Nende intronite RNA moodustab keerulise konserveerunud sekundaarsturuktuuri. See võimaldab neil intronitel olla isesplaissuv. Intronit sisaldav RNA molekul muudab enda kovalentstruktuuri, et täpselt eemaldada intron ning kleepida eksonid omavahel õiges järjekorras kokku. Mõnel juhul osalevad splaissimises ka spetsiaalsed introneid siduvad valgud, mis aitavad introneid voltida isesplaissumiseks vajalikku kolmedimensionaalsesse struktuuri.
Tuuma pre-mRNA intronid (splaissosoomiga eemaldatavad intronid) on ära määratud spetriaalsete intronite järjestustega, mis asuvad eksonite ja intronite vahealadel. Need järjestused tuntakse ära splaissosoomse RNA molekulide abil, kui alustatakse splaissimisreaktsioonidega. Tuuma pre-mRNA intronid on tihti palju pikemad kui neid ümbritsevad eksonid.
Bioloogilised funktsioonid ja evolutsioonEsialgsel hindamisel saab introneid vaadata kui tähtsusetuid järjestusi, mille ainus funktsioon on eemalduda splaissimata algsest RNAst, et valmistada funktsionaalne mRNA, rRNA või tRNA produkt. Siiski, mõned intronid ise kodeerivad spetsiifilisi valke või neid introneid töödeldakse edasi hiljem pärast splaissimist, et genereerida mittekodeerivaid RNA molekule. Alternatiivne splaissimine on laialt kasutusel, et valmistada palju valke ühest geenist, tegemist on post-translatsioonilise geeniregulatsiooniga. Mõned intronid esindavad mobiilseid geneetilisi elemente ja võidakse lugeda kui näideteks isekast DNAst.
Intronite bioloogiline päritolu on selgusetu. Pärast esimest intronite avastust valke kodeerivates geenides eukarüootide rakutuumades, käis arutelu selle üle, kas intronid tänapäevastes organismides on pärit ühisest eellasest või on intronid üsna hiljuti evolutsiooniprotsessis tekkinud. On ka teine teooria: splaissosoom ja intron-ekson struktuurid geenides on jäänuk RNA maailmast. Käib arutelu ka selle üle milline neist hüpoteesidest on õigeim. Viimane populaarne konsensus hetkel on, et intronid on jäänud eukarüootsesse liini isekate elementidena.
Varajased genoomse DNA järjestuse uuringud paljudes organismides on näidanud, et intron-ekson struktuurid homoloogilistes geenides erinevates organismides võivad suuresti varieeruda. Hiljutisemad uuringud kogu eukarüootse genoomi kohta on näidanud, et intronite pikkus ja tihedus varieerub palju sugulasliikide vahel. Näiteks inimese genoom sisaldab keskmiselt umbes 8,4 intronit geeni kohta (139 418 genoomis), üherakuline seen Encephalitozoon cuniculi sisaldab ainult 0,0075 intronit geeni kohta (15 intronit genoomis)). Introneid on evolutsioonis ulatuslikult tekkinud ja/või kadunud evolutsioonis alates eukarüootide ühise eellase lahknemisest. Arvatavasti on see protsess selektsiooni tulem, kalduvusega intronite juurdetekkimisega suuremates liikides nende väiksemate populatsioonide tõttu ja vastupidine efekt toimub väiksemate liikide puhul (eriti üherakulistel liikidel). Bioloogilised faktorid mõjutavad ka millistes geenides intronid kaovad või juurde tekivad.
Intronite alternatiivne splaissimine geenides tekitab suuremat varieetilisust valkude järjestustes, mis transleeritakse ühest geenist, see lubab toota mitmeid erinevaid valke ühest geenist. Alternatiivse RNA splaissimise kontroll teostatakse erinevate signaalmolekulide abil, mis vastavad paljudele rakusisestele ja rakuvälistele signaalidele.
Intronid kui mobiilsed geneetilised elemendidIntronid võivad evolutsioonis minna kaduma või tekkida juude nagu on selgunud paljude ortoloogiliste geenide uuringutest. Selgunud on kaks üldtunnustatud mehhanismi intronite kadumisest, pöördtranskriptaasi-vahendatud intronite kadu (Reverse Transcriptase-Mediated Intron Loss ehk RTMIL) ja genoomideletsioonid. Lõplikud mehhanismid intronite tekkimisest on kirjeldamata ja vastuolulised. Hetkel on teada vähemalt seitse intronite tekkemehhanismi: introni transpositsioon, transposooni insertsioon, tandem genoomi duplikatsioon, introni ülekanne, introni teke homoloogilise rekombinatsiooni tulemusel (DSBR rada), grupp II introni insertsioon ja intronisatsioon.
Introni transpositsiooni korral arvatav introni tekkemehhanism on kui splaissitud intron pöördsplaissub kas enda mRNA või mõne teise mRNA sisse, kus eelnevalt polnud intronit. Intronit sisaldav mRNA seejärel pöördtranskribeeritakse ja tulemuseks on intronit sisaldav cDNA, mis seejärel võib põhjustada intronitekke täieliku või osalise rekombinatsiooni tulemusena oma orignaalgeenilookusesse.
Transposooni insertsioonid võivad ka tulemuseks anda introni tekke. Kui transposoon asetub AGGT järjestusse, duplitseerub see järjestus mõlemal pool transposooni ning selline insertsioon võib introniseerida transposooni, rikkumata koodijärjestust.
Tandemgeeniduplikatsiooni korral doonor- ja aktseprotsplaissimissaitides (mõlemal on AGGT järjestusele sarnane järjestus) eksonisegmendis AGGT järjestuses tandemgeeniduplikatsioon genereerib kaks potentsiaalset splaissumissaiti. Kui splaissosoom selle koha ära tunneb, siis originaalse ja duplitseeritud AGGT vahel olev järjestus splaissitakse geeni koodijärjestust muutmata.
Introni ülekande korral saab pseudogeen või paraloogne geen introni ja siis kannab selle introni üle rekombinatsiooni teel õdeparaloogi, kust intron puudub.
Intronisatsiooni korral tekitavad mutatsioonid uued intronid varasematest eksonijärjestustest. Selle mehhanismi korral ei toimu insertsiooni või DNA generatsiooni.




