Mis on diabeet? Lihtsalt ja selgelt suhkruhaigusest.
Veresuhkruks nimetatakse veres sisalduvat glükoosi. Veres ringlev glükoos on meie keha moodustavatele rakkudele tähtsaim energiaallikas, tagades elutegevuseks vajalikke rakufunktsioonide töö.
Kuidas glükoos verre jõuab?
Süües jõuavad koos muu toiduga meie seedetrakti süsivesikud, mis lõhustatakse glükoosiks. Sealt verre imendudes tõuseb vere glükoosi ehk veresuhkru tase. Seetõttu tõuseb ka tervel inimesel peale söömist veresuhkur. Veresuhkru tõusule reageerib terve inimese kõhunääre insuliini eritamisega, mis viib glükoosi rakku ja veresuhkru mõne aja möödudes endisele tasemele. Veresuhkru taset kontrollitakse organismi poolt rangelt ja erinevate meetoditega. Keha normaalseks toimimiseks ja tervise tagamiseks peab see jääma võrdlemisi kindlatesse piiridesse.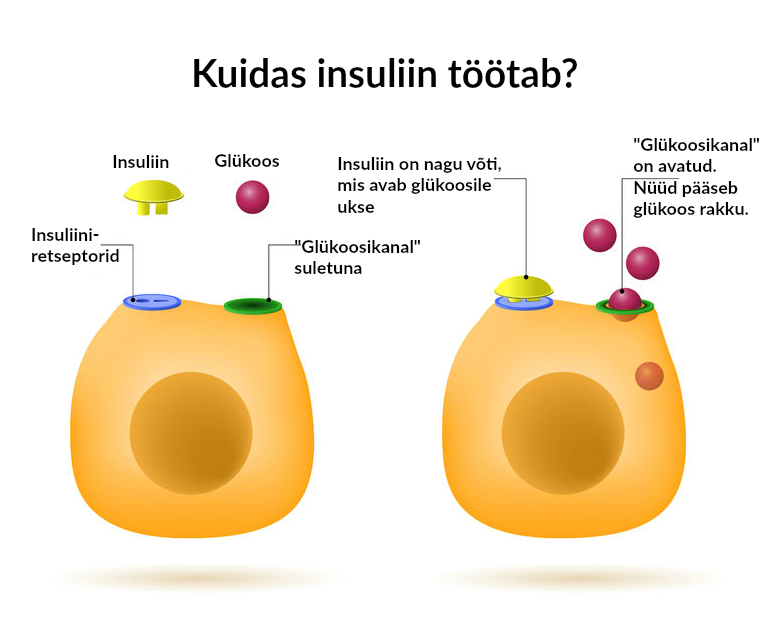
Suhkurtõve korral on insuliini vähe või puudub oma insuliin üldse. Seetõttu glükoosi transport kudedesse on häiritud ja glükoos jääb verre.
Diabeedi ehk suhkruhaiguse saab jaotada kahte põhilisse gruppi:
- tüüpi diabeet
- tüüpi diabeet.
Lisaks on olemas veel vähemlevinud: gestatsioonidiabeet (rasedusaegne diabeet) ning teised suhkrutõve tüübid (pankreatogeenne diabeet, steroiddiabeet, MODY, jt)
Diabeedi kohta kasutatakse ka nime suhkurtõbi (kõnekeeles suhkruhaigus) või suhkurdiabeet ja ladinakeelne nimetus on diabetes mellitus.
1. tüüpi suhkurtõbi tekib enamasti lastel ja noortel, alla 35-aastatel inimestel.
1. tüüpi diabeet kuulub autoimmuunhaiguste hulka. Magusa söömine seda ei põhjusta. Täpne tekkemehhanism on teadmata. Arvatakse, et teatud geneetilise taustaga inimestel tekib keskkonna ja võimalik, et ka viiruste mõjul autoimmuunreaktsioon, mille käigus immuunsüsteem ründab insuliini tootvaid rakke. Koos insuliini tootvate rakkude hävimisega väheneb ka toodetava insuliini hulk, ning kujuneb välja diabeet. Mis paneb immuunsüsteemi valesti käituma, ei ole tänaseni täiesti selge. Sümptomid on janu, sagenenud urineerimine, näljatunne ja samas kaalukaotus, öised jalakrambid, nahasügelus. Diagnoosi hilinemise korral võivad tekkida isutus, iiveldus, kõhuvalu, oksendamine, üldine nõrkus ja ketoatsidoos, isegi teadvusehäired. 1. tüüpi diabeeti haigestunu vajab püsivat ravi insuliiniga, s.t insuliinisüste.
Enamik patsiente ehk 85-90% põeb 2. tüübi diabeeti.
2. tüüpi suhkurtõve põhjuseks arvatakse olevat geneetiliselt determineeritud insuliini sekretsiooni defekt ja häiritud insuliintundlikkus. Enamasti haigestuvad üle 40-aastased inimesed.
2. tüübi diabeedi tekkes on suur roll toitumisel ja keskkonnateguritel ning vähesel füüsilisel koormusel Teist tüüpi diabeeti haigestuvad rohkem vanemad ja 90% osas ülekaalulised inimesed ning ravi alustatakse tablettidega. Insuliinisüsted võivad olla vajalikud aastaid hiljem, kui haigus progresseerub.
Teist tüüpi suhkurtõbi kujuneb välja aegamööda aastate jooksul, esialgu ilma oluliste diabeedi tunnusteta. Veresuhkur tõuseb vähehaaval ja organism kohaneb selle normist kõrgema tasemega. 2 tüüpi suhkruhaigus on salakaval haigus ja haiguse avastamise hetkeks on suhkruhaigus tihti juba organismi kahjustanud.
2. tüüpi diabeet võib avalduda väga erineval moel: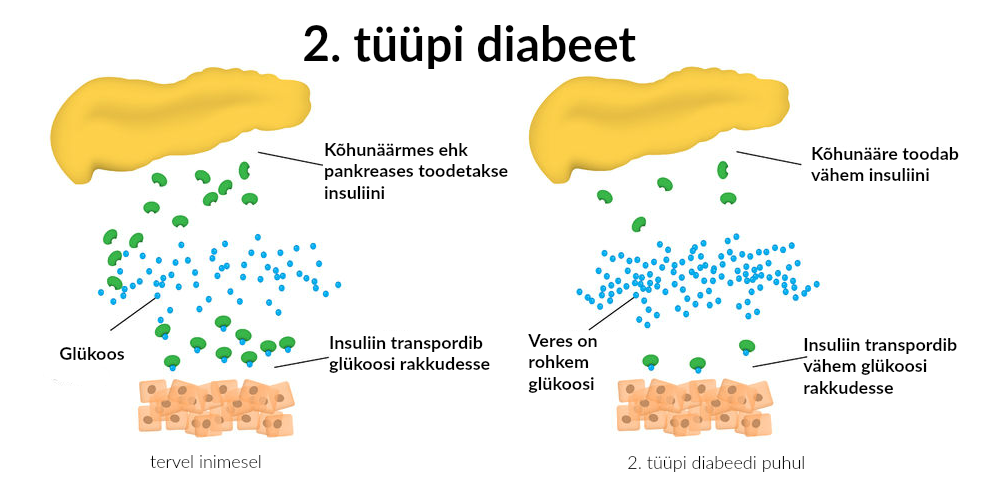
- janu
- sage urineerimine (eriti öösel)
- kaalulangus
- väsimus ja unisus
- suukuivus
- maitse muutused
- nahasügelus
- sagedased mädapõletikud
- tupe ja jalgade seenhaigus
- jalgade valu või tuimuse
- suuõõnehaiguste nagu gingiviit ja parodontoos.
Suhkruhaiguse ravi ja kontrolli all hoidmiseks peab suhkruhaige regulaarselt mõõtma veresuhkrut.
Kui oled juba haigestunud diabeeti, siis peaksid teadma järgmist:
- glükoositase veres kõigub päeva jooksul Seetõttu paljudel juhtudel ei piisa ühest (hommikusest) mõõtmisest päevas. Ära usalda oma enesetunnet, see võib petta.
- veresuhkrut peaks mõõtma mitu korda päevas (loe siit ka vastavat artiklit). Pea nõu oma arsti või diabeediõega, kui tihti ja millal peaks mõõtma;
- hea oleks panna mõõtmistulemused kirja ehk pidada päevikut;
- glükoositaseme kriteeriumid veres on individuaalsed ning sõltuvad mitmest asjaolust (haiguse
 tüüp, vanus jne.).
tüüp, vanus jne.).
Ideaalis, kuid mitte kõigile saavutatav glükoositase peaks tühja kõhuga olema alla 6 mmol/l ning 2 tundi pärast sööki mitte üle 7,8 mmol/l.
Küsi alati oma arstilt, missugune peaks olema sinu glükoositase veres Arsti poolt soovitatud näitudest kinnipidamine tagab töövõime, hea enesetunde ning sul ei ole probleeme tervisega. Diabeedi hea kompensatsioon aitab vältida tüsistuste ehk diabeediga kaasnevate haiguste arengut.








