Ribosoom (inglise ribosome) on nii eel- kui ka päristuumse raku tsütoplasmas esinev kaheosaline molekulaarne masin, mis koosneb ribosomaalse RNA (rRNA) ja valgu molekulidest. Selle ülesanne on katalüüsida peptiidahelate moodustumist, lähtudes raku DNA pealt transkribeeritud informatsiooni-RNA (mRNA, inglise messenger RNA) järjestusest. Seda protsessi nimetatakse translatsiooniks.
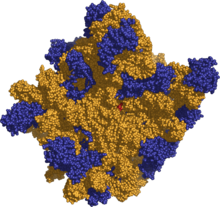
70S ribosoomi 50S alaühik. Kollane osa on rRNA ja sinine märgib valgulist osa. Punasega on märgitud aktiivsait
Samalt valku kodeerivalt DNA järjestuselt võidakse mRNAd sünteesida korduvalt. Eukarüootides toimub seejärel mRNA töötlemine, mille käigus eemaldatakse intronid. Saadud lõpp-produktidele kinnituvad ribosoomid. Nad kasutavad RNA järjestust matriitsina, millele vastavalt seatakse õige koodon vastavusse õige aminohappega. Aminohappeid toovad kohale transport-RNA (tRNA) molekulid, mis sisenevad ribosoomi ja mille antikoodonid paarduvad mRNA järjestusele komplementaarsuse alusel. Ribosoomid on ribosüümid, sest peptiidsideme moodustamine toimub just rRNA abil, millel on peptidüüli transferaasi katalüütiline aktiivsus. Pärast peptiidahela moodustumist pakitakse see funktsionaalseks struktuuriks.
Bakterite, arhede ja eukarüootide ribosoomid erinevad üksteisest oma suuruse, järjestuse, struktuuri ja valk-rRNA suhte poolest. Bakterites võib ühelt mRNA molekulilt transleerida korraga rohkem kui üks ribosoom. Mõned rakuorganellid, näiteks mitokondrid ja kloroplastid, sisaldavad ribosoome, millelt transleeritakse mitmeid just nendes organellides vajaminevaid valke. See on andnud põhjust arvamiseks, et need organellid on evolutsioneerunud algselt eraldiseisvatest prokarüootidest, mis on elama asunud teise raku sisse. Sellest räägib endosümbioositeooria ehk sümbiogenees.
Ribosoom koosneb kahest alaühikust, millest väiksem seondub mRNAga ja suurem tRNA ja ribosoomi saabuvate aminohapetega. Pärast translatsiooni lõppemist jaguneb ribosoom taas kaheks. Prokarüootidel on 70S ribosoomid, mis koosnevad väikesest (30S) ja suurest (50S) alaühikust. Eukarüootidel on 80S ribosoomid, mille alaühikud on vastavalt 40S ja 60S.
Ribosoomide avastajaks võib lugeda George Emil Palade’i, kes kirjeldas 1953. aastal elektronmikroskoobis "tihedaid osakesi või graanuleid". 1974. aastal sai ta koos Albert Claude’i ja Christian de Duvega oma teadustöö eest Nobeli meditsiiniauhinna. 2009. aastal said Venkatraman Ramakrishnan, Thomas A. Steitz ja Ada E. Yonath Nobeli keemiaauhinna ribosoomi struktuuri ja töömehhanismi detailse kirjeldamise eest.
Termini "ribosoom" võttis kasutusele Richard B. Roberts 1958. aastal. 1970. aastate algul püüti terminit eestindada ja pakuti välja "pihukeha", kuid see ei läinud kasutusse ning nii loetakse eesti keeles ainuõigeks tõlkevasteks "ribosoomi".
ÜldiseloomustusRibosoomid koosnevad kahest alaühikust, mis omavahel seondudes moodustavad funktsionaalse organelli. Mõlemad alaühikud koosnevad ühest või mitmest rRNA molekulist ja nendega seondunud valkudest. Kristallograafia abil on näidatud, et valgud asuvad ribosoomi reaktsioonitsentrist väljaspool. See paneb aluse arvamusele, et peptiidsidemete moodustamist katalüüsib RNA. Kirjeldatud teooria kohaselt on ribosomaalsete valkude ülesandeks võimendada RNA sünteesivõimet ja stabiliseerida ribosoomi struktuuri.
Ribosoomid katalüüsivad valkude sünteesi mRNAsse kodeeritud informatsiooni põhjal, kasutades aminohappeid, mille tRNA molekulid on kohale toimetanud. Ribosoomid võivad olla kinnitunud tsütoplasmaatilistele membraanidele, moodustades karedapinnalise endoplasmaatilise retiikulumi, või paikneda tsütoplasmas vabalt.
Ribosoomi alaühikute suuruseid väljendatakse Svedbergi ühikutes (tähis S või Sv), mis iseloomustavad ribosoomi alaühikute sedimentatsioonikiirust tseesiumkloriidi gradiendis tsentrifuugimisel ning sõltuvad nii alaühikute molekulaarmassist kui ka nende kujust.
Prokarüootsed ribosoomid
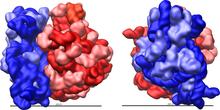
E. coli 70S ribosoom. 50S alaühik on punane ja
30S alaühik sinine. Ribosomaalsed valgud on tähistatud roosa ja helesinise värviga
Prokarüootsete ribosoomide diameeter on ligikaudu 20 nm (200 Å) ja nad koosnevad 65% ulatuses rRNAst ja 35% ulatuses valkudest. Prokarüootidel on 70S ribosoomid, mis koosnevad väikesest (30S) ja suurest (50S) alaühikust. Väikese alaühiku moodustab 16S rRNA (1540 nukleotiidi) ja temaga seondunud 21 valku. Suures alaühikus on 5S rRNA (120 nukleotiidi), 23S rRNA (2900 nukleotiidi) ja 31 valku. Kõiki molekule leidub ühes koopias, välja arvatud valke L12 ja L7. 16S rRNAd saab jagada neljaks ja 23S rRNAd kuueks domeeniks. 5S rRNAd võib lugeda suure alaühiku seitsmendaks domeeniks. Mitokondrid ja kloroplastid, mis on endosümbioositeooria kohaselt evolutsioneerunud bakteritest, sisaldavad samuti 70S ribosoome.
Prokarüootides toimub ribosoomi biogenees tsütoplasmas. Ribosomaalseid valke kodeerib kokku 52 geeni, mis paiknevad 20 operonis. Biogeneesi reguleerib otseselt RNA hulk rakus.
Eukarüootsed ribosoomid
Eukarüootsete ribosoomide diameeter jääb 25 ja 30 nm (250–300 Å) vahele. Nende rRNA-valk suhe on ligikaudu üks. Eukarüootidel on 80S ribosoomid, mis moodustuvad väiksest (40S) ja suurest (60S) alaühikust. Väike alaühik koosneb 18S RNAst (1900 nukleotiidi) ja 33 valgust, suur alaühik 5S RNAst (120 nukleotiidi), 28S RNAst (4700 nukleotiidi), 5,8S RNAst (160 nukleotiidi) ja umbes 49 valgust.
Eukarüootide ribosoomi biogenees toimub üle 200 erineva valgu kaasabil, mis sünteesitakse tsütoplasmas ja transporditakse läbi 120 nm diameetriga pooride tuuma. Ribosomaalne RNA sünteesitakse tuumas piirkonnas, mida nimetatakse tuumakeseks (erandiks on 5S rRNA). Seal seonduvad valgud RNAle ning toimub kompleksi transport tuumast välja. RNAd ei saa üksi transportida, sest ta ei sisalda oma järjestuses vastavat signaali.
Prokarüootide ja eukarüootide ribosoomide erinevusi kasutatakse ära farmakoloogias, et luua antibiootikume, mis pärsiksid bakterinfektsioonide põhjustajate valgusünteesi, kuid ei omaks mingit mõju organismi enda rakkude ribosoomidele. Mitokondrite ribosoomid on kaitstud tänu neid ümbritsevale kahekordsele membraanile, millest antibiootikumid läbi tungida ei suuda.
Asukoht rakusRibosoomid võivad olla kas vabalt tsütoplasmas või kinnitunud endoplasmaatilisele retiikulumile (ER). Mõlemal juhul on aga ribosoomid ise identsed ja nende asukoht sõltub vaid selleks, kas hetkel transleeritava valgu järjestuses on läbi ER transportimiseks vajalik signaal või mitte.
Vabalt tsütoplasmas olevatelt ribosoomidelt sünteesitakse valke, mida kasutatakse hiljem rakus sees. Valgud, mida vajatakse mõnes organellis või mis sekreteeritakse väliskeskkonda, sünteesitakse ERile kinnitunud ribosoomidelt. Sellised valgud liiguvad paralleelselt translatsiooni toimumisega läbi plasmamembraani, kus nad pakitakse vesiikulitesse ja kantakse nende lõpp-sihtkohta. Endoplasmaatilise retiikulumi piirkonda, kuhu sellised ribosoomid on koondunud, nimetatakse "karedaks ERiks".
Funktsioon
Translatsioon
Ribosoomide ülesandeks on transleerida mRNAs peituv informatsioon valgujärjestuseks. mRNA lugemine käib kolmenukleotiidiste koodonite (tripletite) kaupa, millest igale seatakse vastavusse õige aminohape. Igale aminohappele vastab konkreetne tRNA, mis selle ribosoomi transpordib. Aminohappe ja tRNA kompleksi nimetatakse aminoatsüül-tRNAks. Viimane sisaldab oma järjestuses kolmenukleotiidist antikoodonit, mis paardub mRNAl asuva koodoniga komplementaarsuse alusel. Translatsioon toimub suunal 5’ -> 3’.
Valgusüntees algab AUG koodoniga, mis asub mRNA 5’-otsa lähedal. Sellega paardub tRNA molekul, mis transpordib aminohapet metioniin. Prokarüootides on ribosoom võimeline stardikoodonit ära tundma tänu Shine-Dalgarno järjestusele, mis asub sellest 8 nukleotiidi 5’-otsa suunas. Sellega paardub 16S rRNA 3’-otsas leiduv anti-Shine-Dalgarno järjestus. Eukarüootides on sarnane tähtsus Kozaki järjestusel.
Ribosoom sisaldab kolme RNAga seondumise saiti: A, P ja E. A-saidis toimub aminoatsüül-tRNA seondumine. P-saidis sünteesitakse peptiidside. Selle tulemusena jääb tRNA seotuks kogu seni sünteesitud peptiidahelaga. E-saidis seotakse vaba tRNA enne, kui see ribosoomilt vabaneb.
Kahe alaühiku rollid on translatsiooni käigus erinevad. Väike alaühik seondub mRNAga ning sisaldab piirkonda, kus toimub koodoni ja anti-koodoni vaheline interaktsioon. Suur alaühik sisaldab peptidüüli transferaasi tsentrit, kus toimub peptiidsidemete süntees A-saiti seondunud ja P-saiti liikunud tRNAde külge kinnitunud aminohapete vahel ja valmiva peptiidahela hüdrolüüs P-saidi tRNA küljest translatsiooni lõppfaasis. Peptidüüli transferaasi tsentri juurest algab ka polüpeptiidide väljutamiseks mõeldud tunnel, mida mööda valguahelad saavad stabiilselt läbi väikese alaühiku liikuda.






