Embrüogenees ehk organismi looteline areng on ontogeneesi esimene staadium. See algab munaraku viljastumisest ja lõpeb munast koorumisega (lindudel ja enamikul muudel loomadel), sünnimomendiga (imetajatel ja muudel elussünnitajatel loomadel) või idu moodustumisega seemnes (taimedel). Embrüogeneesi võib jaotada varaseks ja hiliseks embrüogeneesiks. Inimese puhul lõpeb varajane embrüogenees siis, kui embrüo on arenenud looteks.

Inimese embrüogenees
Eri liikide loodete omavaheline võrdlus näitab, et imetaja embrüo sarnaneb algselt kala lootega, seejärel kahepaikse ja roomaja omaga ning alles lõpuks omandab imetajale omased tunnused. Seega läbitakse ontogeneesi alguses (embrüogeneesis) liigi evolutsioonilise arengu ehk fülogeneesi etapid.
Viljastumine ja sügoot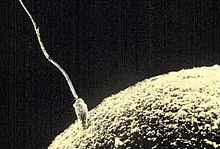
Sperm viljastamas munarakku
Munarakk on asümmeetriline, tal on animaalne poolus (tulevane ektoderm ja mesoderm) ja vegetatiivne poolus (tulevane endoderm). Munarakk on kaetud teda kaitsvate kihtidega. Esimene kiht on vitelliinmembraan (imetajatel zona pellucida), mis on seotud munaraku rakumembraaniga ja koosneb glükoproteiinidest. Erinevates taksonites esinevad erinevad rakulised ja mitterakulised kihid, mis ümbritsevad vitelliinmembraani.
Viljastumine on isas- ja emasgenoomi konjugatsioon kahe gameedi liitumise tagajärjel. Tänu kahe gameedi liitumisele taastatakse keharakkudele omane diploidne kromosoomistik ning aktiveeritakse munarakk arenema. Loomadel toimub protsessi käigus spermatosoidi ühinemine munarakuga, mis viib embrüo arenema. Sõltuvalt loomaliigist võib viljastumine toimuda kas kehasiseselt või kehaväliselt. Viljastatud munarakku nimetatakse sügoodiks.
Lõigustumine ja moorula
Lõigustumine ja gastrulatsioon
Lõigustumine on loomade (sh ka inimese) viljastunud munaraku ehk sügoodi kiire mitootiline jagunemine, ilma et tütarrakud vahepeal kasvaksid. Lõigustumine lõpeb kobarloote ehk moorula moodustumisega. Lõigustumise eesmärgiks on organismi hulkraksuse taastamine ja rakutuuma-tsütoplasma suhte normaliseerumine. Kui keharakkude tuuma-tsütoplasma suhe on tavaliselt 1:10, siis spermil on see suhe 1:1, munaraku puhul aga kaldub tugevasti tsütoplasma kasuks, 1:1000 või veelgi rohkem. Lõigustumine on jagunemine ilma kasvamiseta, mis on vajalik tuuma-tsütoplasma suhte normaliseerimiseks väga tsütoplasmarikka munaraku baasil. Lõigustumisel moodustuvaid tütarrakke nimetatakse blastomeerideks. Lõigustumine võib olla sünkroonne või asünkroonne, st blastomeerid võivad jaguneda kas üheaegselt või teevad seda eri ajal. Enamikul loomarühmadel on lõigustumise algul sünkroonse lõigustumise periood, mis on mõõdetav 4–14 rakutsükliga ning läheb seejärel üle asünkroonseks perioodiks. Ainult imetajatel ei ole sünkroonset lõigustumist ja esimesed kaks blastomeeri alustavad kohe lõigustumist asünkroonselt. Intensiivne paljunemine, ajal nagu see viljastatud munaraku moorulaks kujunemine on, toimub nendesamade geenide abil, mis vähkkasvaja rakkude vohamist põhjustavad.
Lõigustumise tüübid
Lõigustumise tüüp sõltub rebu hulgast. Lõigustumine võib olla täielik ehk totaalne või osaline ehk partsiaalne.
- Täielik – vähese või keskmise rebuhulgaga ootsüüdid.
- Täielik lõigustumine jaguneb:
- totaal-ekvaalne ehk isoletsitaalne (blastomeerid ühe suurusega),
- totaal-adekvaalne ehk mesoletsitaalne (animaalse pooluse blastomeerid veidi väiksemad) lõigustumine (käsnad, keriloomad, mõned vähid, süstikkala ja imetajad),
- totaal-inekvaalne: blastomeerid on märgatavalt erineva suurusega – mikromeerid ja makromeerid (kahepaiksed, kopskala, kammloomad).
- Osaline – suure rebuhulgaga.
- Osaline lõigustumine jaguneb:
Gastrulatsioon
Gastrulatsioon toimub siis, kui ühekihiline
blastula voltub sissepoole, suureneb ja moodustub
gastrula. Värvikood: ektoderm – sinine; endoderm – roheline; blastotsööl – kollane; arhenteron – lilla.
Gastrulatsiooni käigus paigutuvad lõigustumisel tekkinud rakud ümber oma õigetele kohtadele, pannakse paika mitmekihilise organismi kehaplaan ja toimub tüvirakkude determinatsioon ehk programmeerimine. Morfoloogiliselt iseloomustab gastrulatsiooni loote- ehk idulehtede moodustumine.
Lootelehti on kolm: ektoderm ehk välisleht, entoderm ehk siseleht ja mesoderm ehk keskleht. Hüdralaadsetel on vaid ekto- ja endoderm.
Gastrulatsioon toimub pärast lõigustumist ning blastula ja ürgjuti moodustumist. Gastrulatsioonile järgneb organogenees, mille käigus arenevad lootelehtedest elundid ehk organid.
Kõikidel loomaliikidel moodustuvad samadest lootelehtedest samad elundid ja elundkonnad. Need muutused toimuvad järk-järgult embrüonaalse induktsiooni alusel. Kuna rakud vahetavad informatsiooni teiste ümbritsevate rakkudega, siis ühtede rakkude diferentseerumine viib ka vastavalt teiste rakkude diferentseerumiseni. Näiteks närvitoru moodustumine mõjutab mesodermi teket. Viimane omakorda soodustab närvitoru arenemist kesknärvisüsteemiks (pea-ja seljaajuks). Seega määrab ühtede kudede või elundite teke teiste kudede ja elundite tekke.
Gastrulatsiooni molekulaarne mehhanism ja ajastus on organismiti varieeruv, kuid triploblastiliste organismide hulgas esinevad mõned üleüldised sarnasused:
- muutused embrüo topoloogilises struktuuris: kerakujulisest lihtsalt ühendatud pinnalt keeruliselt ühendatud pinnaks,
- rakkude diferentseerumine üheks kolmest tüübist (ento-, meso- või ektodermiks),
- suur osa entodermaalseid rakke omab seedefunktsiooni.
OrganogeneesNärvisüsteemi areng

Neuraalplaadi sulgumine neuraaltoruks
Närvisüsteem on esimene elundkond, mis hakkab arenema pärast gastrulatsiooni. Blastotsööli sisserullunud kordomesoderm indutseerib enda kohal ektodermis neuraalplaadi. Neuraalplaat süveneb esialgu neuraalvaoks ja sulgub seejärel neuraaltoruks. Närvitoru morfogeneesis on tähtis osa nii sisemistel (rakusisestel) kui ka välistel teguritel. Sisemisteks teguriteks, mis kujundavad närvitoru, on mikrotorukeste ja mikrofilamentide lokaalsed iseärasused. Esimeseks tõukeks närvivao kujunemisel on muutused ventraalsetes mediaansetes rakkudes, mis on kontaktis seljakeelikuga. Need rakud moodustavad närvivao mediaanse hinge (analoogia aknahingega). Sealsed rakud pikenevad esialgu mikrotorukeste paralleelsete kimpude koondumise abil ja seejärel omandavad nad kiilu kuju tipmiste rõngasjalt paiknevate mikrofilamentide kontraktsiooni tulemusel. Järgmine etapp närvitoru kujunemisel on lateraalsetes hingedes kiilukujuliste rakkude moodustumine. Kogu kesknärvisüsteemi edasine arenemine on seotud rakkude lokaalse vohamisega ja valikulise hukkumisega. Selle tulemusena moodustub närvitoru eesosas esialgu kolm ja seejärel viis ajupõiekest. Kolme ajupõiekese staadiumis on eristatavad eesaju (prosencephalon), keskaju (mesencephalon) ja rombaju (rhombencephalon). Eesaju jaguneb edaspidi kaheks: otsajuks (telencephalon) ja vaheajuks (diencephalon). Rombaju jaguneb omakorda tagaajuks (metencephalon) ja piklikajuks (myelencephalon s. medulla oblongata).
Neuraalhari
Neuraalplaadi piirkonnad, mis lähevad üle ektodermiks, on tulevase neuraalharja allikaks. Neuraalhari on selgroogsetel niivõrd tähtis, et teda nimetatakse neljandaks looteleheks. Neuraalharjast pärineb hulk rakutüüpe. Neuraalharja võib jagada neljaks piirkonnaks:
- Pea neuraalhari, mille rakkudest areneb kraniofatsiaalne mesenhüüm, mis annab pea- ja näopiirkonna kõhred, luud ja muu sidekoe. Need rakud võtavad osa neelutaskute seinast ja annavad alguse tüümusele, hamba odontoblastidele ja sisekõrva kõhrele.
- Kere neuraalhari, millest pärinevad pigmenti sünteesivad melanotsüüdid. Samuti arenevad nendest dorsaal- ja sümpaatilised ganglionid, neerupealise säsiosa ning närvikogumikud ümber aordi.
- Ristluu piirkonna neuraalhari, mille rakkudest genereeritakse soole parasümpaatilised ganglionid.
- Kardiaalne neuraalhari, mis paikneb pea ja kere neuraalharja vahel kana embrüos 1. kuni 3. somiidini ja moodustab suurte arterite lihaselise seina ja kopsutüve septumi.
Neuraalharja rakud rändavad oma kohtadele mööda dorsaalset ja ventraalset trajektoori. Dorsaalne tee läheb läbi somiitide anterioorse osa, mis on kindlaks tehtud mitmesuguste markeritega. Need rakud annavad sensoorsed ja sümpaatilised neuronid, neerupealise säsi ja Schwanni rakud.
Seede- ja hingamiselundite kujunemine
Endoderm moodustab seedetrakti epiteliaalse voodri. Sooltoru jaguneb kolmeks osaks: ees-, kesk- ja tagasooleks. Eessool areneb kõige kiiremini ja moodustab oma lateraalses osas neelu- ehk lõpustaskud, millest imetajatel on selgelt eristatavad neli ja lindudel viis paari. Esimesest paarist lõpustaskutest kujunevad kuulmetõrved, teine paar lõpustaskuid kasvab kinni, täitub lümfaatilise koega ja moodustab kurgumandlid. Kolmanda ja neljanda lõpustasku dorsaalsed osad arenevad tüümuseks ja ventraalsed osad neljaks kõrvalkilpnäärme sagaraks. Viimased liituvad kilpnäärmega, mille alge sopistub välja kolmanda ja neljanda neelutasku paari vahel. Eessoole tagaosast sopistub bronhiaaljätke, millest arenevad kopsud. Kesksool jääb lindudel ja imetajatel ühendusse rebukotiga. Maks areneb kesksoolest ühe jätke kujul ventraalselt ja pankreas kahe jätke kujul nii kõhtmisest kui ka selgmisest piirkonnast. Tagasoolest sopistub välja allantois ja lindudel veel kloakaal- ehk Fabriciuse paun. Viimane on lümfaatiline elund. Imetajatel on homoloogiline kude hajutatud piki jämesoole seinu. Suu- ja anaalpiirkonnas puutub ektoderm kokku endodermiga, millest arenevad vastavalt suu- ja anaalava. Hüdra näitel on leitud, et tegelikult jääb ekto- ja endodermi vahele ühekihiline eriline rakkude populatsioon, mis suuava avanemisel venituvad ilma ühegi raku purunemiseta. Suuavast tungib ektoderm dorsaalselt Rathke tasku näol vaheaju ja endodermi vahele ja sellest piirkonnast areneb hüpofüüsi eessagar.
Mesodermi liigestumine
Neurula staadiumis jaguneb mesoderm viieks regiooniks:
- Kordomesoderm, mis formeerib seljakeeliku ja see määrab kogu organismi anterioposterioorse telje.
- Somiitide dorsaalne mesoderm annab organismi seljapiirkonna mesodermaalsed koed.
- Vahelmine ehk intermediaarne mesoderm, millest areneb kuse- ja suguelundkond.
- Külgplaatide mesoderm, mis annab alguse südamele, veresoontele ja vererakkudele, kõhu- ja rinnaõõnsuste sisevoodrile ning jäsemete mesodermaalsetele komponentidele, v.a lihased.
- Pea mesoderm, millest arenevad näo sidekoed ja muskulatuur.
Paraksiaalne mesoderm segmenteerub somiitideks. Somiit jaguneb edasises arenemises kolmeks osaks: selgmiseks dermatoomiks ja kõhtmisteks sklerotoomiks ja müotoomiks. Dermatoomist areneb pärisnahk, sklerotoomist aksiaaltoes ja müotoomist seljalihased. Vahelmisest mesodermist arenevad neerude üksikud põlvkonnad. Inimesel 22.-st ja hiirel kaheksandast arengupäevast pärast viljastumist areneb esimeste somiitide kõrval vahelmisest mesodermist pronefrose juha, millest imetajatel kujunevad umbsed pronefrose torukesed. Alamatel selgroogsetel avanevad pronefrose torukesed tsöloomi. Nende läheduses tsöloomi seinas areneb tihe veresoonevõrgustik kompaktseks päsmaks. Nendest veresoontest surutakse välja vereplasma ilma valkudeta ja pronefrose torukestes imendub enamik vereplasmat tagasi. Lämmastiku ainevahetuse lõpp-produktid, üleliigsed soolad ja vesi väljutatakse pronefrose juha kaudu. Pronefrose juha jaguneb posterioorselt kaheks: mesonefrose ehk Wolffi juhaks ja paramesonefrose ehk Mülleri juhaks. Mesonefrose torukesed paiknevad üksikutes segmentides sümmeetriliselt ja on seotud individuaalsete veresoonte päsmakestega. Isastel loomadel säilib osa mesonefrose torukestest ja moodustavad munandi somaatilise karkassi, munandimanuse, Wolffi juhast kujuneb seemnejuha. Emastel mesonefrose torukesed ja Wolffi juha taandarenevad ning Mülleri juhast arenevad munajuhad ja emakas. Amniootide püsineer on metanefros, mis areneb mesonefrosest kaudaalsemalt. Wolffi juhast sopistub kusejuha pung, mis hargneb tugevasti mesodermis (“ureeter-puu”) ja indutseerib seal neerutorukeste morfogeneesi. Neerutorukesed lõpevad umbsete kihnudena, mis ümbritsevad veresoonte päsmakesi ja koos moodustavad neerukehakesed. Kusejuha pungast areneb neeruvaagen. Kogu metanefrose arengus on tähtis koht neuraalharja rakkudel ja adhesioonimolekulidest sündekaanil.









