Blogi, mis räägib kõigest, mis on Leonhardile oluline ja/või huvitav. Kommenteerige, tellige, lugege, nautige ja õppige.
Sissejuhatus
Tänapäeval ei kujutaks me ette geneetikat ilma DNA uuringuteta. Geneetilised uuringud on muutunud nii lihtsaks ja kättesaadavaks, et igal inimesel on võimalik saata enda DNA laborisse, et saada rohkem teada oma päritolu kohta või uurida pärilike haiguste eelsoodumuse kohta. Kuid kas teadsite, et geenide pärandumise seaduspärad avastati ligikaudu sada aastat enne seda, kui mõisteti DNA rolli päriliku info salvestamisel?
Õpieesmärgid
Selle peatüki lõpuks:
- kirjeldate Mendeli katsetes ilmnenud seaduspärasid;
- seostate Mendeli katsetes ilmnenud seaduspärasid nende geneetilise põhjendusega;
- lahendate Mendeli seadustel põhinevaid ülesandeid;
- selgitate Mendeli seaduste rakendusvõimalusi.
Mendeli katsed
Esimesed avastused pärilikkuse seaduspärade kohta tegi Tšehhi munk ja kooliõpetaja Gregor Mendel juba 19. sajandi keskel. Enne Mendeli avastusi usuti, et vanemate tunnused segunevad järglastes sarnaselt nagu sinine ja kollane värv annavad segades rohelise. Mendel arvas seevastu, et pärilikkus tuleneb teatud muutumatute pärilikkusüksuste edasi andmisest. Tema teooria kohaselt on organismis igast tunnusest kaks versiooni, mille organism saab oma vanematelt. Oma teooria katsetamiseks ristas ta kloostriaias herneid ning uuris tunnuste pärandumist (joonis 3.3.5.1.). Pärilikkuse uurimist selliste meetoditega, mida kasutati enne DNA rolli avastamist, nimetatakse tänapäeval klassikaliseks geneetikaks.
Joonis 3.3.5.1. Gregor Mendel selgitas hernetaimi kasvatades ja uurides välja põhilised tunnuste pärandumise seaduspärad.
Mendeli katsed hernestega
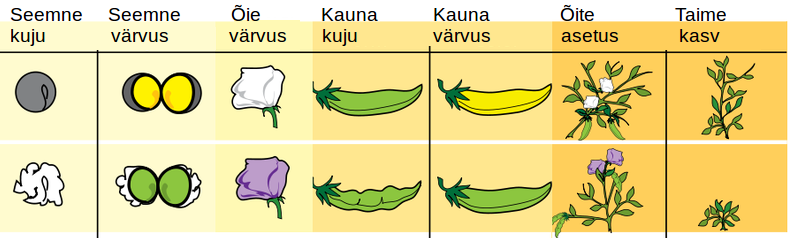
Mendel uuris oma katsetes seitsme tunnuse pärandumist: herneseemnete värv ja kuju, kauna värv ja kuju, õite värv ja asetus ning taime kasv. Esialgu ristas ta taimi, mis erinesid omavahel ainult ühe tunnuse poolest. Esmalt kasvatas ta eri tunnustega taimi eraldi peenardel. Seeläbi selgitas ta välja, et ühest puhtast sordist taimed annavad alati täpselt samasuguseid järglasi, näiteks rohelise herne taimedelt saab alati roheliste seemnetega järglasi (joonis 3.3.5.2.).
Joonis 3.3.5.2. Mendel ristas erinevate tunnustega herneid ning uuris tunnuste pärandumist.
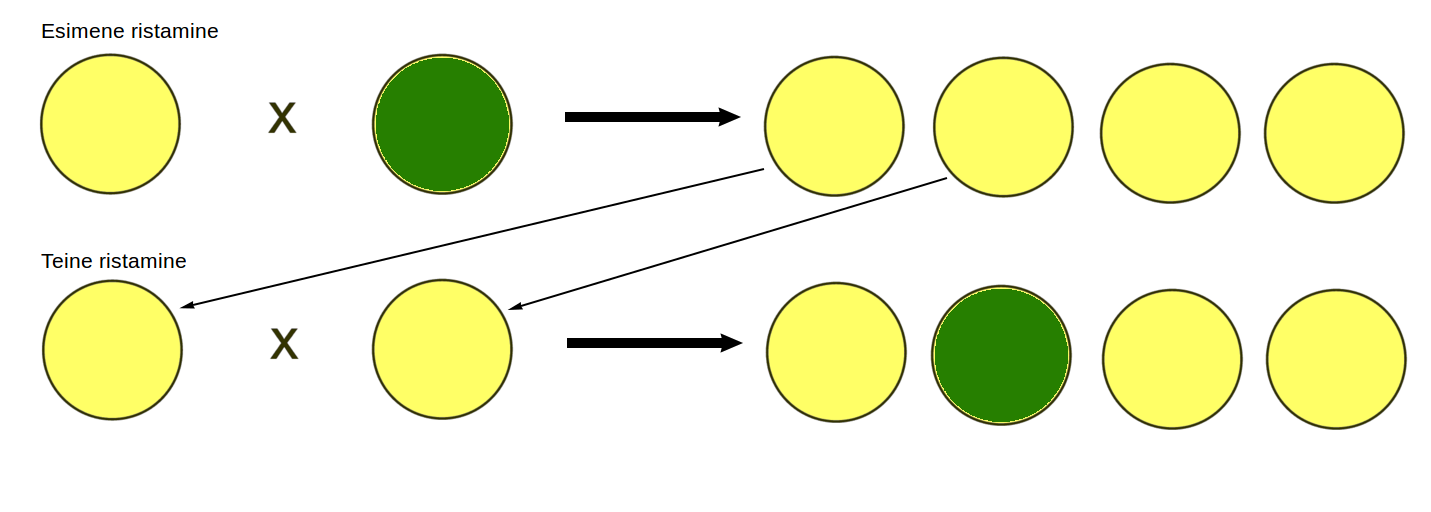
Seejärel ristas Mendel omavahel erinevaid hernesorte, näiteks kollaseid herneid rohelistega. Katse tulemusena selgus, et igast tunnuste paarist leidus üks domineeriv tunnus, mis pärast ristamist saadud järglastes avaldus. Puhtasse sorti kuuluvate kollaste ja roheliste herneste ristamisel saadi ainult kollaseid herneid, krobelisi ja siledaid herneid ristates oli tulemuseks aga vaid siledad herned (joonis 3.3.5.3.).
Joonis 3.3.5.3. Mendel avastas oma katsete käigus, et igas tunnustepaaris leidus üks domineeriv tunnus.
Ristates esimese järglaste põlvkonna herneid omavahel sai Mendel aga hoopis teistsugused tulemused. Roheliste ja kollaste herneste ristamisel saadud kollaseid herneid omavahel uuesti ristates olid järglastest ¾ kollased ning ¼ rohelised (joonis 3.3.5.4.). Sarnased osakaalud olid iseloomulikud kõigi vaadeldud tunnuste puhul. Sellest järeldas Mendel, et teises põlvkonnas tunnused lahknevad uuesti.
Joonis 3.3.5.4. Kollaste ja roheliste herneste ristamisel saadud järglased on kõik ühetaoliselt kollased. Saadud kollaseid herneid omavahel ristates lahknevad tunnused uuesti ning 1/4 saadud hernestest on rohelised.
Simulatsiooni abil saate ka ise Mendeli katseid läbi viia proovida. Selleks vajutage esmalt "Breed" ehk rista, et luua kaks juhuslikku vanemat, ja uuesti "Breed", et näha nende vanemate järglasi. Seejärel valige kaks järglast, keda soovite omavahel ristata, ning vajutage taaskord "Breed", et näha järgmise põlvkonna järglasi. Saate protsessi korrata nii mitu korda kui soovite, kuid 6 põlvkonna järel hakkavad vanemad põlvkonnad kaduma. Kui soovite alustada simulatsiooni uuesti uute juhuslike vanematega, siis uuendage lehte või sulgege see ja avage uuesti. Kas oskate simulatsiooni abiga leida samad seaduspärad, mis Mendel oma katsetega tuvastas?
Mendeli seadused
Tänapäeval saame Mendeli avastusi põhjendada teadmistega geenide toimimisest. Teame, et organismi keharakkudes on kahekordne kromosoomistik ning igast geenist on kaks versiooni, millest üks pärineb emalt, teine isalt. Selliseid geenide versioone nimetatakse alleelideks. Kui organismil on mingi geeni mõlemad alleelid ühesugused, nimetatakse seda homosügootsuseks, kui erinevad, siis heterosügootsuseks. Mendeli esimesed katsed olid sordipuhaste taimedega, mis andsid ainult ühesuguste tunnustega järglasi, järelikult oli seal tegemist homosügootsete ehk samasuguste alleelidega taimedega.
Erinevate tunnustega organismide ristamise tulemusi on kõige parem selgitada skeemi abil. Kokkuleppeliselt tähistatakse sellisel skeemil puhtast sordist vanemaid tähega P (ladina keeles parentes ehk vanemad) ning nende järglasi tähega F (ladina keeles filii ehk lapsed). Seda, mitmenda põlvkonna järglastega on tegemist, tähistatakse numbriga, näiteks F1 ja F2.
Mendeli järgmistes katsetes oli erinevateks alleelideks näiteks seemne kollane või roheline värvus. Tähistades kollast värvust määrava alleeli A tähega ning rohelist värvust määrava alleeli a tähega, ilmneb, et puhtast sordist kollaste herneste alleelid on AA ja roheliste herneste alleelid aa (joonis 3.3.5.5.). Sugurakkude valmimisel jääb igasse sugurakku üks alleel. See tähendab, et kollaste herneste sugurakkudesse jäävad alleelid A ja A, roheliste herneste sugurakkudesse a ja a. Kollase ja rohelise herne sugurakkude liitumisel on seega ainsad võimalikud kombinatsioonid, mis saavad tekkida, Aa, ehk hernes, kus on üks kollast ning üks rohelist värvust määrav alleel.

Joonis 3.3.5.5. Kollaste ja roheliste puhtast sordist herneste ristamisel on tulemuseks herned, millel on üks kollast, üks rohelist värvust määrav geenivariant. Kuna kollane on domineeriv tunnus, siis on välimuselt kõik herned kollased.
Hoolimata sellest, et kõigil F1 põlvkonna hernestel on mõlemad alleelid, on kõik herned väliselt ühetaolised ehk kollased. Järelikult avaldab värvusele mõju ainult üks alleelidest. Seda alleeli, mis ristamise järel avaldub, nimetatakse dominantseks alleeliks, ja teist alleeli nimetatakse retsessiivseks. F1 põlvkonna järglastel on kõigil üks dominantne ja üks retsessiivne alleel ehk nad on heterosügootsed ning kõik järglased on omavahel identsed. Seda järeldust nimetatakse Mendeli esimeseks seaduseks.
Vaatame nüüd skeemil, mis juhtub, kui ristata omavahel F1 põlvkonna heterosügootseid järglasi, kellel on Aa alleelid ehk üks kollast, teine rohelist värvust määrav alleel. Sugurakkude valmimisel jääb kummalgi hernel osadesse sugurakkudesse kollast (A) ja teistesse rohelist (a) värvust määrav alleel. Ühe taime sugurakud liituvad teise taime sugurakkudega, järelikult on uues ehk F2 põlvkonnas võimalik kolm erinevat kombinatsiooni: AA, Aa ning aa (joonis 3.3.5.6.). Kuna domineeriv tunnus ehk kollane värvus avaldub nii AA kui ka Aa kombinatsiooni korral, retsessiivne tunnus ehk roheline värvus aga vaid aa kombinatsiooni korral, on lihtne näha, miks ¾ järglastest on domineeriva ning ¼ retsessiivse tunnusega. Nende tulemuste põhjal on sõnastatud Mendeli teine seadus: F2 põlvkonnas lahknevad tunnused kindlates suhetes.
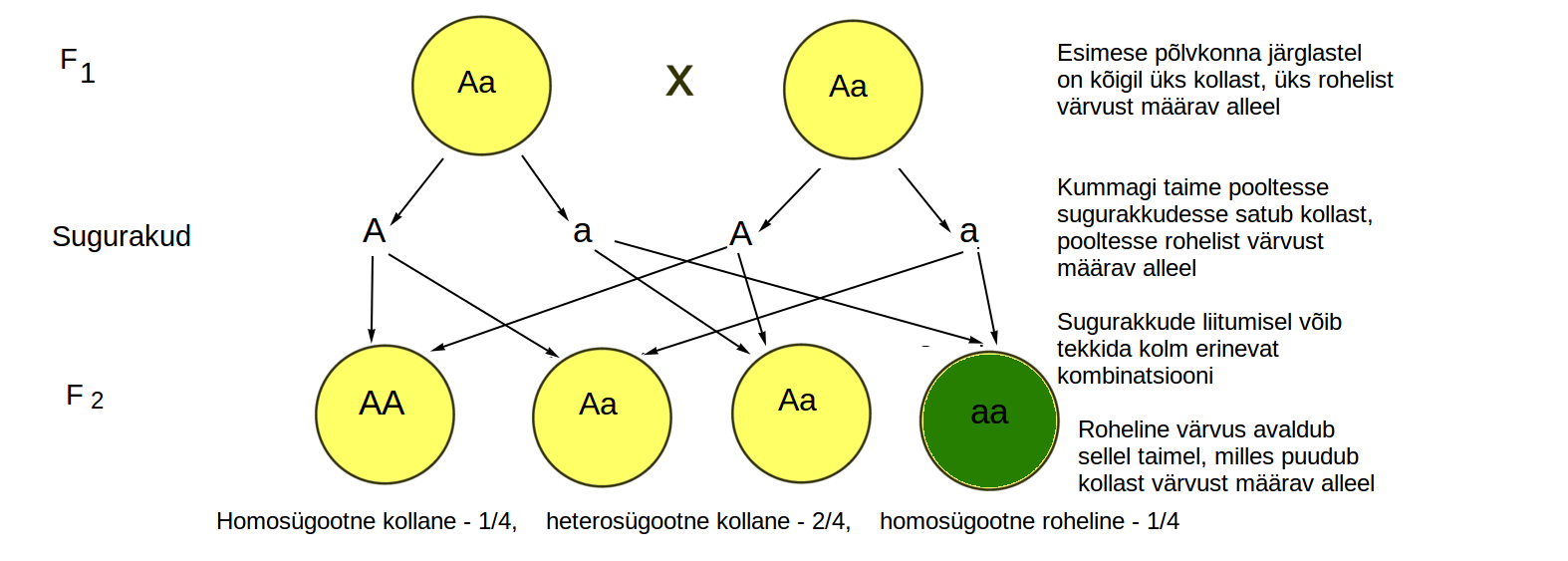
Joonis 3.3.5.6. F1 põlvkonna ristamisel omavahel lahknevad tunnused kindlas vahekorras. 3/4 järglastest avaldub dominantne tunnus ning 1/4 retsessiivne tunnus.
Uurides mitme erineva tunnusega herneste lahknemist järeldas Mendel ka seda, et tunnused päranduvad üksteisest sõltumatult. Seda seaduspära nimetatakse Mendeli kolmandaks seaduseks. Tänapäeval teame, et see seadus kehtib ainult tunnuste puhul, mida määravad geenid asuvad erinevates kromosoomides või samas kromosoomis, kuid üksteisest kaugel. Lähedal asuvad geenid päranduvad koos, kuna need antakse sama kromosoomiga järglastele edasi. Seetõttu avalduvad osad tunnused enamasti koos.
Mendeli seaduste rakendusvõimalused
Kuigi Mendeli katsed tuginesid herneste sihipärasele ristamisele, on tema avastatud seadustel palju laiem rakendus. Mendeli seadused kehtivad ka looduses juhuslikult sündivate järglaste puhul, sealhulgas inimestel. Samas tuleb silmas pidada, et mitte kõik tunnused ei pärandu vastavalt Mendeli seadustele. Inimestel on puhtaid Mendeli seaduste kohaselt päranduvaid tunnuseid küllaltki vähe. Paljud tunnustest on määratud mitme geeni poolt. Selliseid tunnuseid nimetatakse polügeenseteks. Polügeensed tunnused on näiteks silmade värv, kasv, kehakaal ja nahavärv (joonis 3.3.5.7.).
Joonis 3.3.5.7. Kasv ja silmavärv on polügeensed tunnused ehk nende tunnuste määramisel osaleb rohkem kui üks geen.
Enamikul tunnustest ei suru üks alleel teist täielikult maha. Olukorda, kus heterosügoodil avaldub kahe homosügoodi vahepealne tunnus, nimetatakse intermediaarsuseks. Näiteks saab sageli punaste ja valgete õitega lillede ristamisel roosade õitega järglased. Mõnikord võivad aga mõlema alleeli poolt määratud tunnused avalduda samaaegselt. Seda nimetatakse kodominantsuseks ning siis saadakse punaste ja valgete õitega lillede ristamisel hoopis punase-valge-kirjud lilled (joonis 3.3.5.8.).
Joonis 3.3.5.8. Kui punaste ja valgete õitega taimede ristamisel saadakse roosade õitega taim, on tegemist intermediaarsusega, kui aga punase-valgekirjud õied, siis kodominantsusega.
Mendeli seadused on siiski olulised, kuna võimaldavad leida mitmete haiguste, näiteks daltonismi ehk punarohepimeduse (joonis 3.3.5.9.) ja hemofiilia ehk vere hüübimatuse esinemise tõenäosust järglastel. Selleks on vaja teada, kas vanemad on selle tunnuse suhtes homo- või heterosügootsed. Katseloomade- ja taimede puhul ristatakse selleks dominantsete tunnustega isendit retsessiivsete tunnustega isendiga. Kui järglased on kõik ühesugused, on tegemist homosügootsusega, kui aga toimub lahknemine suhtes 1:3, siis on tegemist heterosügootse vanemaga. Inimeste puhul loomulikult selliseid meetodeid kasutada ei saa, seetõttu tuleb lasta DNA-d analüüsida või uurida oma sugupuud.
Joonis 3.3.5.9. Daltonismi ehk puna-rohe-pimesuse korral ei suuda inimene eristada punaseid ja rohelisi toone. Sellisel juhul ei ole inimene võimeline pildil olevat numbrit lugema.
Kokkuvõte
Keharakkudes on igast geenist kaks versiooni ehk alleeli, mis mõjutavad tunnuse avaldumist. Dominatne tunnus avaldub ka heterosügootidel, retsessiivne tunnus ainult homosügootidel. Mendeli seaduste kohaselt päranduvad tunnused üksteisest sõltumatult ning kindla seaduspärasuse alusel. Homosügootsete vanemate ristamisel on F1 põlvkonna järglased kõik heterosügootsed ning ühetaolised, F2 põlvkonna järglastel toimub aga tunnuste lahknemine. Kõik tunnused ei pärandu Mendeli seaduste kohaselt, kuid Mendeli seaduste tundmine aitab mõista mitmete haiguste pärandumist ning arvutada haigete järglaste saamise tõenäosust.
Mõisted
alleel
homosügootsus
heterosügootsus
dominantsus
retsessiivsus
polügeensus
intermediaarsus
kodominantsus

 [Public domain], via Wikimedia Commons")