AB0 süsteem (AB0 system) -- inimese vererühmade AB0-süsteem, esimene tunnustesüsteem, mille puhul tõestati looduslik geneetiline polümorfism ja polüalleeelsus. Selle vererühmasüsteemi avastas seroloogilise uurimisega Austria arst K. Landsteiner 1900. a. Geneetiline tingitus ja struktuur avastati 1910-20ndatel aastatel. Meditsiinilises käsitluses eristatakse 4 AB0-rühma (nn. veregruppi): 0, A, B ja AB. Need on määratud ühe geeni 3 alleeliga, millest 2 (IA ja IB) determineerivad erinevaid glükosiidseid antigeene erütrotsüütide pinnal, 3. alleel (i e. I0) on aga funktsioonita. A- ja B-alleelid on teineteise suhtes kodominantsed, 0-alleeli suhtes aga dominantsed. 6 genotüüpi määravad vererühmi (fenotüüpe) järgmiselt: IAIA ja IAi - A, IBIB ja IBi - B, IAIB - AB ning ii (e. I0I0) - 0. Tegelikult on A-tüüp heterogeenne: on eristatavad alleelid IA1 ja IA2 ning vastavalt fenotüübid A1, A2, A1B ja A2B. Seega 4 alleeli, 10 genotüüpi ja 6 fenotüüpi (vererühma). (Alleel IA2 on IA1 suhtes retsessiivne, kuid teiste alleelidega avaldub samamoodi kui IA1). Viimast (A1-A2) eristust kasutatakse peamiselt populatsioonigeneetikas. AB0 süsteemi polümorfismis esinevad etnilised ja rassilised erinevused alleelide ja seega genotüüpide ning fenotüüpide sagedustes. AB0-geeni kromosoomne lokalisatsioon on 9q31.3. Inimesegeneetika moodsa nomenklatuuri kohaselt on selle geeni sümboliks AB0 ja alleelid tähistatakse: AB0*A1, AB0*A2, AB0*B, AB0*0.
aditiivne geenitoime (additive gene action) -- vt. polümeerne geenitoime.
aheldus (linkage) -- vt. geeniaheldus, geneetiline aheldus.
ahelduskaart (linkage map) -- kromosoomi (või selle osa) geneetiline kaart, millele on kantud selles sisalduvad lookused aheldusanalüüsi põhjal leitud lineaarses järjestuses ja nendevaheliste geneetiliste kaugustega (mõõtühik: cM = sentimorgan).
aheldusrühm (linkage group) -- aheldusastme alusel lineaarselt järjestatud geneetiliste lookuste (eelkõige geenide) rühm mingis pidevas geneetilises strutuuris (kromosoomis, plasmiidis jms.). Aheldusrühma kaugemate elementide vaheline aheldus (rekombinatsioonisageduse piiratus) järk-järgult nõrgeneb, kusjuures rühma eri otste lookused ei pruugi olla omavahel geneetiliselt aheldunud (st. rekombineeruvad vabalt). Tuumalookuste täielike aheldusrühmade arv võrdub kromosoomide arvuga genoomis. Aheldusrühma skeemi, millele on kantud lookuste nimed ja nendevahelised geneetilised kaugused sentimorganites (cM), nimetatakse geneetiliseks kaardiks.
alleel (allele) -- geeniteisend, geeni esinemisvorm; üks kahest või mitmest alternatiivsest geenivariandist, mis asuvad populatsiooni isendite homoloogiliste kromosoomide samas lookuses ja toimivad sama tunnuse kujunemisele (tekitades selle eri vorme või avaldumisastmeid. Viimasel ajal kasutatakse alleeli mõistet laiemas tähenduses: mis tahes genoomse lookuse (nt. restriktsioonisaidi, minisatelliidi jm.) alternatiivne vorm. Meioosis alleelid lahknevad eri rakkudesse, st. välistavad teineteist. (Vt. ka alleelsus).
alleelide interaktsioon (allele interaction) -- alleelide koostoime tunnuse kujundamisel, mis väljendub alleelide fenogeneetilises avaldumises heterosügootse genotüübi puhul. Eristatakse kolme tüüpi alleelide interaktsiooni: dominantsus (vastaspoolusena retsessiivsus), kodominantsus ja intermediaarsus.
alleelsus (allelism) -- geenide (ja muude lookuste) omadus esineda erinevate teisenditena - alleelidena. Alleelid tekivad üksteisest geenmutatsioonide tagajärjel. Geenide alleelsus põhjustab tunnuste erinevate (alternatiivsete) variantide (või avaldumistasemete) olemasolu perekonna/populatsiooni indiviididel (kuid erinevad alleelid võivad avalduda ka ühtmoodi). Mingil geenil võib populatsioonis esineda alleele üks (monomorfsus), kaks (dialleelsus) või rohkem (polüalleelsus). Haploidse organismi või haplofaasi (gameedi, eose) genotüübis (haplotüübis) on iga geen esindatud mingi ühe alleeliga; diploidse organismi või diplofaasi (sügoodi) geenipaarid võivad olla homosügootsed ja heterosügootsed. Polüalleelsuse korral on diploidsete organismide populatsioonis võimalike genotüüpide arv (antud geeni suhtes) 1/2k(k+1), kus k on alleelide arv seerias. Leviku ja kohasuse alusel eristatakse normaalalleele (mets- e. ulukalleele, wild alleles) ja (haruldasi) mutantalleele (defektalleele, letaalaleele), fenogeneetilisevastasmõju järgi heterosügootses genotüübis dominantseid, retsessiivseid, kodominantseid, mittetäielikult dominantseid ja intermediaarseid alleele.
alloploidsus (alloploidy) -- hübriidse indiviidi (ka somaatilise rakuhübriidi) kromosoomistiku koosnevus eriliigilistest kromosoomikomplektidest. Viimased võivad esineda ühekordselt (allodiploidsus e. amfihaploidsus) või mitmekordselt (allopolüploidsus). Allodiploidsed organismid on tavaliselt viljatud (kromosoomide meiootilise paardumise puue). Taimeliikide vaheliste hübriidide kromosoomistiku kahekordistumine (allotetraploidsus e. amfidiploidsus) tagab tihti normaalse meioosi ja viljakuse. Vrd. autoploidsus.
aneuploidsus (aneuploidy) -- kromosoomistiku ebavõrdne kordsus indiviidi rakkudes, mõne kromosoomi väiksem või suurem arv võrreldes kromosoomistiku üldise kordsusega; kõrvalekalle euploidsusest. Aneuploidsus võib esineda igasuguse ploidsusastme korral; selle eristamiseks lisatakse vastava euploidse kordsuse nimetusele liited hüpo- või hüper- (nt hüperhaploid, hüpodiploid, hüpertriploid jne). Diploidsete organismide (nt. inimese) aneuploidsuse tähistamiseks kasut. ka järgmisi mõisteid ja sümboleid nullisoomik (2n-2), monosoomik (2n-1), trisoomik (2n+1), kaksiktrisoomik (2n+1+1), tetrasoomik (2n+2). Aneuploidsust põhjustavad homoloogiliste kromosoomide või tütarkromatiidide lahknemise häired meioosis või (harvem) mitoosis.
asutaja efekt (founder effect) -- e. rajaja efekt, geneetilise triivi üks äärmuslikke erijuhte, mis tuleneb populatsiooni asutamisest (e. rajamisest) väga väikese arvu isendite poolt, äärmisel juhul üksikisendeist, kes on pärit mingist suurest populatsioonist. Asutajate väikese arvu tõttu on uue populatsiooni geenifond juhuslikel (valimivea) põhjustel oluliselt erinev emapopulatsiooni omast: selles puuduvad paljud alleelid, osa tavalisi alleele võib saada haruldaseks ja, vastupidi, mõni haruldane alleel võib saada sagedaks ning võivad fikseeruda ka mõned uued mutatsioonid. Asutaja efekt ilmneb peamiselt juhul, kui uus asutatud populatsioon on emapopulatsioonist täielikult või oluliselt isoleeritud -- nt. ookeanisaarte, eraldatud järvede, mäestikuorgude, uute mandrite jne. koloniseerimisel. Kuna asutatud populatsioon on vähemalt algperioodil vähesearvuline, siis toimib seal tugevalt ka tavaline (kestev e. persistentne) geneetiline triiv. Nende asjaolude tõttu võib uus populatsioon täiesti juhuslikel põhjustel saada uue evolutsioonisuuna kuni uue alamliigi, liigi või perekonna tekkeni välja.
autoploidsus (autoploidy) -- samaliigiliste kromosoomikomplektide kordsus (autodiploidsus, autopolüploidsus) indiviidi rakkudes. Kõik kromosoomikomplektid on üksteisele homoloogsed ja paarisploidsuse (di-, tetra- jne) korral paarduvad meioosis normaalselt. Kasutatakse vastandmõistena alloploidsusele. Vt. ka ploidsus.
autosoom (autosome) -- kromosoom, mis esineb võrdarvuliselt (nt paariliselt) liigi kõigil indiviididel, olenemata nende soost. Inimesel on 22 autosoomi, mida tähistatakse numbritega 1...22 (kahanevas pikkusjärjestuses). Vrd. gonosoom.
autosoomsus (autosomy) -- geenide ja muude markerite lokaliseeritus autosoomides; neist geenidest määratud tunnuste päritavusviis. Vastavaid geene ja tunnuseid nim. autosoomseteks. Autosoomsete (autosomal) geenide pärandumine ja kombineerumine toimub klassikaliste Mendeli seaduste järgi: nad päranduvad mõlemalt sugupoolelt ühtmoodi kõigile järglastele (retsiprooksed ristamised on võrdväärsed). Aautosoomsuse vastandnähtus on gonosoomsus ehk suguliitelisus.
Barri kehake (Barr body) -- vt. sugukromatiin.
biomeetriline geneetika (biometrical genetics) -- e. kvantitatiivne geneetika (quantitative genetics), geneetika haru, mis uurib (pideva) kvantitatiivse muutlikkusega tunnuste päritavust. Kvantitatiivne muutlikkus (erinevused tunnuse avaldumisastmes) põhineb mitmete (tihti paljude) geenide polümeersel e. aditiivsel koostoimel ja samaaegsel keskkonnatingimuste mõjul. Tunnuse pideva muutlikkuse korral ei ole võimalik tuvastada ei osalevaid geene ega individuaalseid genotüüpe. Põhiprobleemiks on selliste (mõõdetavate) tunnuste muutlikkuse dispersioonikomponentide lahutamine ja geneetiliselt tingitud osa hindamine populatsioonis, st. päritavuskoefitsiendi arvutamine. Selleks kasutatavad meetodid seisnevad erineva sugulusastmega indiviidide tunnuse rühmasisese dispersiooni või korrelatsiooni võrdlemises. Inimese puhul kasutatakse kaksikute, perekonnastatistilist ja adopteeritute uurimise meetodit. Biomeetrilise geneetika alusepanija oli inglise teadlane Francis Galton oma töödega (alates 1865. a.) inimese vaimsete võimete ja iseloomuomaduste päritavuse alalt.
C-meioos (C-meiosis) -- meioosi anomaalne vorm, mis on tingitud kolhitsiini (colchicine) või muu käävimürgi toimest. Olenevalt sellest, kas käävi moodustumine on pärsitud ühes või mõlemas meiootilises jagunemises, tekivad diploidsel organismil kas diploidsed või tetraploidsed rakud (gameedid või spoorid).
C-mitoos (C-mitosis) -- mitoosi anomaalne vorm, mis on tingitud kolhitsiini (colchicine) või muu käävimürgi toimest; mitoos katkeb metafaasis. Soodsal juhul tekitab emarakuga võrreldes kahekordse kromosoomistikuga tütarrakud. Praktikas kasutatakse peamiselt polüploidsete taimesortide saamiseks.
C-väärtus (C-value, constant-value ) -- DNA konstantne kogus eukarüootse organismi genoomi (kromosoomikomplekti) kohta. Diploidses rakutuumas on DNA koguse väärtus võrdne 2C-ga. Jagunevate diploidsete rakkude S-perioodi lõpus ning G2-perioodis ja mitoosi (meioos I) pro- ning metafaasis on C-väärtus 4C. Haploidsed rakud (seega diploidsete organismide spermid ja munarakud) sisaldavad 1C DNA-d. Eukarüootide C-väärtus varieerub väga ebakorrapärasel viisil: paljudel evolutsiooniliselt alamate (st. morfoloogiliselt ja füsioloogiliselt keerukuselt lihtsamate) organismide C-väärtus on mitmeid (isegi kümneid) kordi suurem, kui evolutsiooniliselt kõrgematel (hilisematel) vormidel. See nähtus on tuntud C-väärtuse paradoksina. Nüüd on teada, et sellised erinevused on tingitud mitte-geense (st. mittekodeeriva) DNA hulga suurest varieeruvusest erinevates fülogeneetilistes liinides.
dialleelsus (diallelism) -- uuritava geeni esinemine kahe alleeli kujul populatsiooni (liigi) geenifondis.
diploidsus (diploidy) -- 1) liigiomase kromosoomikomplekti kahekordsus indiviidi (raku) kromosoomistikus (2x). 2) liigi normaalsele diplofaasile (sügootidele) vastava kromosoomistiku olemasolu (2n). Valdaval enamikul loomadel langevad mõlemad määratlused kokku; polüploidsetel liikidel (paljudel taimedel) on diplofaasis kromosoomikomplektide arv üle kahe (2n > 2x). Vt ploidsus, haploidsus, polüploidsus.
dominantsus (dominance) -- domineerimine, ühe tunnuse (alleeli) prevaleerumine tunnusepaaris (alleelipaaris) teise üle; heterosügootse genotüübi (Aa) avaldumine samamoodi dominantse homosügootse genotüübiga (AA); selliselt prevaleerivat tunnust (alleeli) nim. dominantseks. Polüalleelsuse korral võib esineda astmeline dominantsus (A¹>A²>A³). Dominantsus ja retsessiivsus on ühe nähtuse eri poolused. Meditsiinigeneetikas nim. dominantseks mutantset alleeli (defekti, haigust), mis avaldub heterosügootse genotüübi puhul (mutantne homosügoot võib avalduda tegelikult teisiti -- raskemal kujul või letaalsena).
ekspressiivsus (expressivity) -- mingi genotüübi (enamasti mutatntse alleeli) fenotüübilise avaldumise viis ja tase; on iseloomustatav nii kvalitatiivsest kui ka kvantitatiivsest küljest. Nagu penetrantsuski, sõltub see nii genotüübilisest tagapõhjast kui ka keskkonnatingimustest, sh. ka indiviidide soolistest erinevustest.
endomitoos (endomitosis) -- somaatilise polüploideerumise vorm, toimub tavaliselt diferentseerumise lõppfaasi rakkudes. See on mitoosi sarnane protsess, mis kulgeb intaktses tuumas ja ilma raku jagunemiseta ning tekitab polüploidse raku. Endomitoosi korral kromosoomid kahestuvad ja kondenseeruvad nagu normaalses mitoosiski, kuid tuumakate jääb püsima ja kääviaparaati ega metafaasiplaati ei moodustu; endoanafaasis eralduvad kromatiidid autonoomselt; seejärel läheb tuum jälle interfaasi seisundisse. Endomitoos on üks polüsomaatia põhjusi.
endoreduplikatsioon (endoreduplication) -- e. varjatud endomitoos (masked endomitosis), kromosoomide kahestumine ja kromosoomistiku kordistumine interfaasi tuumas ilma mingite mitoosile omaste vaadeldavate ilminguteta. Endoreduplikatsioon on kõige levinum somaatilise polüploideerumise ja polüsomaatia tekke protsess. Rakkude korduv endoreduplikatsioon võib tekitada kõrgpolüploidseid rakke. Endoreduplikatsiooni erivorm on polütenisatsioon, mille puhul korduvalt kahestunud kromatiidid ei eraldu ning tekivad polüteensed kromosoomid.
epistaas (epistasis) -- ühe geeni tõkestav, pärssiv või varjutav toime teise lookuse geeni avaldumisele. Geeni, mis allutab teise geeni avaldumise, nimetatakse epistaatiliseks (E,e), alluvat geeni hüpostaatiliseks(H,h). Kui epistaatilise geeni ainsaks avaldumisviisiks on teise geeni pärssimine, siis nimetatakse teda inhibiitor- või supressorgeeniks. Eristatakse dominantset (E>H,h) ja retsessiivset (ee>H,h) epistaasi. Epistaasi erijuhtudel võib dihübriidsel ristamisel F2-põlvkonnas saada fenotüübilisi lahknemisi (ühe tunnuse variantide) sagedussuhetes 9:3:4 (retsessiivne epistaas), 12:3:1 või 13:3 (dominantne epistaas). Biomeetrilises geneetikas nimetatakse igasugust mitteaditiivset geenide interaktsiooni epistaasiks.
eukarüoot (eukaryote) -- organism, kelle rakud on päristuumset (eukarüootset) tüüpi. Kõik taimed, seened ja loomad on eukarüoodid. Enamus eukarüoote on hulkraksed (koelis-organilise ehitusega) ja tõelise (holomiktilise) sugulise sigimisega.
eukarüootne rakk (eukaryotic cell) -- e. päristuumne rakk; evolutsiooniliselt kõrgema ja keerukama organisatsiooniga rakutüüp, vastandatav prokarüütsele rakule. Võrreldes prokarüootse rakuga on eukarüootne rakk suuremate mõõtmetega; tema peamised geneetilised struktuurid asuvad membraanidega piiristatud tuumas; genoom on jaotunud mitmeks (vähemalt kaheks) kromosoomiks, milles DNA on püsivalt seotud histoonvalkudega; tsütoplasmas on rohkesti membraanseid organelle (mitokondrid, tsütoplasmavõrgustik, Golgi kompleks, lüsosoomid, taimerakkudes plastiidid, vakuoolid); rakujagunemine põhineb keerukatel kromosoomide jaotamise ja kombineerimise mehhanismidel (mitoos või meioos).
eukromatiin (euchromatin) -- vt. kromatiin.
euploidsus (euploidy) -- kromosoomistiku võrdkordsus, genoomi kõigi kromosoomide (erandiks võivad olla sugukromosoomid) võrdne arv indiviidil (rakus). Vastandmõiste on aneuploidsus.
feen (phene) -- geneetiliselt kontrollitav elementaartunnus; fenotüübi väikseim osis, mis võib individuaalselt muutuda mingi geenmutatsiooni tagajärjel ja kombineeruda teiste feenidega.
fenogenees (phenogenesis) -- geneetiliselt kontrollitud tunnuste (feenide) kujunemiskäik indiviidi ontogeneesis; molekulaarsete, biokeemiliste, füsioloogilste, morfoloogiliste jm. protsesside ning muundumiste jada geenist tunnuseni. Fenogeneesis ilmneb alleelide ja erinevate geenide interaktsioon või ka ühe geeni erisuunaline toime erinevatele tunnustele (polüfeensus).
fenokoopia (phenocopy) -- fenotüübi mittepärilik, mingist keskkonnategurist põhjustatud modifikatsiooniline muutus, mis sarnaneb teatud geenmutatsioonist tingitud muutusega.
fenotüüp (phenotype) -- indiviidi (morfoloogiliste, füsioloogiliste, keemiliste, etoloogiliste, arenguliste) tunnuste (variantide ja avaldumistasemete) vaadeldav kogum; kitsamas mõistes üksiku uuritava tunnuse ilmetüüp (variant, seisund). Fenotüüp kujuneb indiviidi arengus (fenogeneesis) genotüübis sisalduva info realiseerumise tulemusena, tihti sõltuvalt elukeskkonna tingimustest.
fenotüübiline muutlikkus (phenotypic variability) -- individuaalsete ja/või populatsiooniliste erinevuste olemasolu fenotüüpides, mis võib olla tingitud kas genotüübilistest erinevustest (vt. geneetiline muutlikkus) või modifikatsioonidest (vt. modifikatsiooniline muutlikkus).
geen (gene) -- kromosoomi kindlas lookuses paiknev pärivustegur, mis määrab otse või kaudselt (tihti koostoimes teiste geenidega) ühe või mitme tunnuse arengu; DNA-molekuli funktsionaalne lõik, mis tavaliselt sisaldab informatsiooni (mRNA vahendusel) ühe valgu (või polüpeptiidi) sünteesiks (kuid on ka nt rRNA ja tRNA geenid, mis valgumolekule ei kodeeri). Eukarüootide geenid on enamasti katkendliku struktuuriga: kodeerivate nukleotiidijärjestuste (eksoonide) vahel on mittekodeerivad järjestused (introonid); geeni pidevus mRNAs on tagatud pre-mRNA töötlusega tuumas (RNA protsessing, splaissing).
Eristatakse struktuur- ja regulaatorgeene. Geenid eksisteerivad struktuurilt ja tihti ka funktsioonilt erinevate alleelide kujul.
geeniaheldus (gene linkage) -- geneetilise ahelduse kitsam, klassikaline mõiste, mis tähendab piiratud (mittevaba) rekombineerumist geenide vahel genotüüpide pärandumisel, st. Mendeli III (sõltumatu lahknemise) seaduse rikkumist. Geeniaheldus on tingitud geenilookuste lähestikusest paiknemisest piki kromosoomi. Aheldunud geenid rekombineeruvad ristsiirde kaudu, mille sagedus oleneb geenidevahelisest kaugusest kromosoomis (Morgani seadus).
geenide duplikaatsus (multiplikaatsus) (duplicate or multiplicate gene action) -- kordne geenitoime, kaks või mitu dominantset (mittealleelset) geeni kujundavad nii eraldi kui ka ühistoimes ühe ja sama dominantse tunnuse; ainult homosügoot mõlema või kõigi geenide retsessiivsete alleelide suhtes annab alternatiivse (retsessiivse) tunnuse (nt. A-B- = A-bb = aaB- > aabb). Geenide duplikaatsus avaldub tunnuste (ka haiguste) geneetilise heterogeensusena, kus mitmete eri geenide mutatsioonid põhjustavad fenotüübiliselt samu tunnusevariante (haigusi). Kahe duplikaatse geeni korral ilmneb F2-põlvkonnas fenotüübiline lahknemine kahe variandi vahel suhtes 15:1.
geenide interaktsioon (gene interaction) -- kahe või enama (mittealleelse) geeni koostoime ühiselt määratava tunnuse fenogeneesis. Enamasti eristatakse järgmisi geenide iinteraktsiooni tüüpe: komplementaarsus, epistaas, duplikaatsus (või multiplikaatsus) ja polümeersus e. aditiivsus.
geenide komplementaarsus (gene complementary) -- geenide vastastikune täiendavus; kahe või enama mittealleelse geeni teatud (tavaliselt dominantsete) alleelide (A-B-) koostoimel tekib uus tunnus, mis nende geenide eralditoimel (aaB-, A-bb) ei esine. Kahe paari komplementaarsete geenide poolest erinevate homosügootide ristamisel on F1-põlvkonna isenditel kummagi vanema tunnusest erinev uus tunnus; F2-põlvkonnas võib esineda, olenevalt komplementaarse geenitoime erijuhtudest, fenotüübilisi lahknemisi ühe tunnuse 2...4 variandi vahel sagedussuhetes 9:7, 9:3:4, 9:6:1, 9:3:3:1.
geenidoos (gene dosage) -- geeni (alleeli) efektiivsete koopiate arv indiviidi genotüübis. Geenidoosi erinevused indiviidide vahel võivad olla tingitud ploidsuse erinevustest (vt. euploidsus ja aneuploidsus), homo- ja heterosügootsusest, suguliitelisusest (homosügootsus vs. hemisügootsus), genotüübilistest erinevustest polümeerse geenitoime korral. Nt. genotüüpidevahelised fenotüübilised erinevused alleelide intermediaarse vastasmõju korral on tingitud geenidoosist. Suguliiteliste geenide puhul toimivad mitmesugused geenidoosi kompensatsiooni mehhanismid.
geenidoosi kompensatsioon (gene dosage compensation) -- geenide avaldumise regulatsioonimehhanism, mis tagab geenide võrdväärse avaldumise X-liiteliste geenide puhul homo- ja heterogameetsel sugupoolel. Erinevatel organismirühmadel on vastav mehhanism erinev, kõige uuritum on see imetajatel. M. Lyon (1966) avastas, et emasimetajate somaatiliste rakkude kahest X-kromosoomist inaktiveeritakse üks varases embrüogeneesis fakultatiivse heterokromatinisatsiooni (kondensatsiooni) teel (rakutuumas tekib Barri kehake e. sugukromatiin). See inaktivatsioon on igas rakus homoloogiliste X-kromosoomide suhtes juhuslik, nii et emasisendi kudede erinevates rakkudes talitlevad erinevad homoloogid (seda nähtust nim. laionisatsiooniks). Selline mehhanism tagab seda, et emasorganismi rakkudes talitleb samamoodi üks koopia X-liitelisi geene, nagu hemisügootses isasorganismis, kuid emasisend on talitlevate suguliiteliste geenide suhtes mosaiikne -- erinevates rakukloonides avalduvad eri alleelid. Osa X-kromosoomi lühikese õla tipuosa geene ei allu laionisatsioonile; nende doosikompensatsiooni tagavad spetsiaalsed regulaatorgeenid (vt. Xg-süsteem). Generatiivsetes rakkudes (gonotsüütides, ovotsüütides) laionisatsiooni ei toimu.
geenifond (gene pool) -- e. genofond, populatsiooni kõigi isendite genotüüpides olevate geenide (alleelide) ja muude geneetiliste elementide kogum. Geenifondi iseloomustab iga geeni puhul olemasolevate alleelide arv ja nende sagedused ning vastava geeni suhtes võimalike genotüüpide arv ja nende sagedused.
geenitriiv (gene drift) -- vt. geneetiline triiv.
geenmutatsioon (gene mutation) -- pärilik muutus mingi geeni molekulaarses struktuuris. Geenmutatsioonide tagajärjel tekivad geenide teisendid, alleelid. Geeni lineaarse struktuuri erinevates kohtades toimuvaid mutatsioone, mis võivad omavahel rekombineeruda, nim. punktmutatsioonideks. Geenmutatsioonid võivad olla fenotüübiliselt avalduvad või mitteavalduvad [sünonüümsed koodonimuutused, mittekodeerivate piirkondade (introonide) muutused, asendused kodeerivates piirkondades, mis avalduvad RNA/valgu primaarstruktuuri muutustena, kuid ei põhjusta muutusi nende molekulide funktsionaalses (ruumilises) struktuuris, retsessiivsed mutatsioonid heterosügootses genotüübis]. Avalduvad geenmutatsioonid on sageli kahjulikud (nt. põhjustavad inimese pärilikke haigusi ja muid puudeid).
genealoogia (genealogy) - indiviidi suguvõsalisse põlvnemisse puutuv, selle uurimine sugupuu koostamise abil. Inimesegeneetikas kasutatakse genealoogilist meetodit tunnuste (defektide, haiguste) päriliku sõltuvuse ja päritavusviisi (auto- või gonosoomsus, dominantsus või retsessiivsus, geneetiline aheldus) ning üksikindiviidide genotüübi selgitamiseks.
geneetiline aheldus (genetic linkage) -- geneetiliste markerite (geenide ja muude järjestusmarkerite) seostatus pärandumisel, mis piirab nende vaba kombineerumist e. sõltumatut lahknemist. (st. nad päranduvad valdavalt vanematele omastes alleelsetes kombinatsioonides). Geneetiline aheldus on tingitud geneetiliste markerite lähestikusest paiknemisest ühes lineaarses struktuuris, ühes kromosoomis või muus genoomses nukleiinhappemolekulis (NH). Geneetilised markerid, mis paiknevad ühes kromosoomis (ühes NHs) moodustavd ühe aheldusrühma. Geneetiline aheldus. avaldub selles, et diheterosügoot AB/ab moodustab suurema sagedusega vanemtüüpi gameete (P = AB + ab) kui rekombinantseid (R = Ab + aB), st. f = (P1+P2) /(R1+R2) > 1.
geneetiline kaart (genetic map) -- e. ahelduskaart (linkage map)
geneetiline lahknemine (genetic segregation) -- 1) alleelide eraldumine paaridest ja jaotumine erinevate rakkude (gameetide, eoste) ning neist arenevate indiviidide vahel. Selle põhjuseks on homoloogiliste kromosoomide paaride lahknemine meioosis. Geneetiline lahknemine on geneetilise analüüsiga tuvastatav üksnes heterosügootsete geenipaaride puhul, ta on Mendeli lahknemisseaduste olemuseks. 2) Geneetiliselt erinevate indiviidide esinemine heterosügootide (hübriidide) järglaskonnas vastavalt Mendeli lahknemisseadustele või Morgani aheldusseadusele. See lahknemine on meioosi ja viljastumise koostoime tagajärg. Eristatakse genotüübilist ja fenotüübilist lahknemist. Erinevad fenotüübilise lahknemise viisid samal genotüübilise lahknemise alusel on tingitud alleelide ja mittealleelsete geenide interaktsiooni tüüpidest, mõnikord ka modifikatsioonilisest muutlikkusest.
geneetiline muutlikkus (genetic variability) -- pärilik (heritable) muutlikkus, erinevuste teke või esinemine biosüsteemide (indiviidide, rakkude, organellide, viiruste, populatsioonide) vahel, mis on tingitud muutustest nende geneetilistes struktuurides (genotüübis, genoomis, geenifondis). Geneetilise muutlikkuse põhjusteks on mutagenees (mutatiivne muutlikkus) ja geneetiline rekombinatsioon (kombinatiivne muutlikkus). Avaldumistaseme järgi eristatakse genotüübilist ja fenotüübilist muutlikkust (suur osa genotüübilist muutlikkust on varjatud, st. ei avaldu fenotüübiliste erinevustena).
geneetiline polümorfism (genetic polymorphism) -- mingi lookuse kahe või enamaalleeli (ja tihti ka fenotüübi) esinemine populatsioonis, kusjuures ka haruldasima alleeli sagedus on suurem, kui vahetu mutatsiooniline teke seda võimaldaks. Kasutatakse mitmeid tinglikke polümorfismi kriteeriume, kuid enamasti rakendatakse 1%-kriteeriumi, mille kohaselt defineeritakse lookus (geen) polümorfseks, kui selles on vähemalt kaks alleeli harvima sagedusega vähemalt 1%. Eristatakse varjatud ja avalduvat polümorfismi, üleminevat (e. ajutist), adaptiivset ja neutraalset polümorfismi ning regionaalset polümorfismi.
geneetiline rekombinatsioon (genetic recombination) -- eri isendite geneetiliste elementide ümberkombineerumine järglaste uue geneetilise struktuuriga (alleelse koosseisuga) kromosoomideks, haplo- ja genotüüpideks. Geneetiline rekombinatsioon põhjustab indiviidide kombinatiivse muutlikkuse. See on tüüpiline suguliselt sigivate eukarüüotsete organismide pärilikkusemehhanismile, kus meioos ja viljastumine kombineerivad vanemate genoomielemente geneetiliselt erinevatesse sigirakkudesse (gameetidesse või eostesse) ja sügootidesse. Eristatakse interkromosoomset rekombinatsiooni (eri kromosoomide ja neis sisalduvate geenide sõltumatust lahknemisest tulenev) ja intrakromosoomset rekombinatsiooni (ühes kromosoomipaaris paiknevate geenide ümberkombineerumine homoloogide vahel ristsiirde kaudu). Prokarüootidel ja viirustel esineb mitmesuguseid osalise geneetilise rekombinatsiooni (paraseksuaalseid) protsesse (nt. seksduktsioon, transduktsioon, transformatsioon).
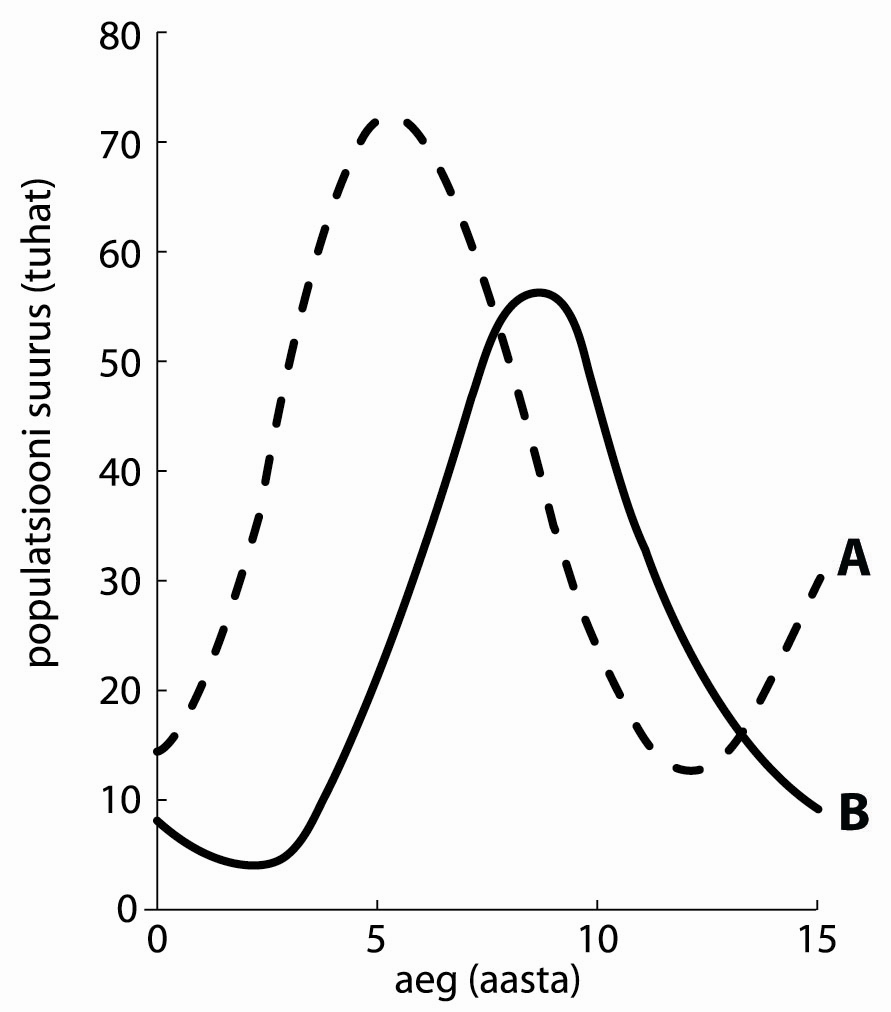
geneetiline triiv (genetic drift, random genetic drift) -- geeni alleelide (ja mittegeensete geneetiliste markerite alleelsete variantide) sageduse juhuslikud muutused populatsiooni järjestikustes põlvkondades. Geneetiline triiv on oma statistiliselt olemuselt valimiviga alleelide ülekandel vanempõlvkonna geenifondist järglaspõlvkonna omasse ja oleneb seega populatsiooni efektiivsest suurusest: ta on seda intensiivsem, mida väiksem on populatsioon. Alleelisageduse muutuste võimalikku amplituudi põlvkonna kohta saab hinnata tema dispersiooni kaudu: s2 = p*(1-p)/(2Ne), kus p on alleelisagedus vanempõlvkonnas ja Ne on populatsiooni efektiivne suurus. Geneetiline triiv mõjutab eelkõige valikuliselt neutraalsete või nõrga valikusurve all olevate alleelide sagedusi, kusjuures tema toime lõpptagajärjeks (kui ta toimib piisavalt kaua) on ühe alleeli fikseerumine geenifondis ja teiste eliminatsioon, st. ta viib populatsiooni geneetilise muutlikkuse vähenemisele ja kaole. Väga väikestes populatsioonides on geneetilise triivi efekt sarnane inbriidingu toimele. Geneetiline triiv antud populatsioonis võib olla kestev (persistent drift) või ajutine e. mööduv (intermittent drift), viimase erivormideks on asutaja efekt ja pudelikaela efekt. Nüüdisaegses evolutsiooniteoorias peetakse geneetilist triivi tähtsaks evolutsiooniteguriks, mis on põhjustanud geenide, populatsioonide ja liikide (aga võib-olla ka kõrgemat järku taksonite) mitteadaptiivseid muutusi ja erinevusi.
genoom (genome) -- ühes liigiomases kromosoomikomplektis sisalduv geneetiline materjal. Genoomi iseloomustatakse kromosoomide (resp. DNA molekulide) arvu ja tüüpide, DNA-koguse (summaarse molekulmassi või nukleotiidipaaride arvu), DNA nukleotiidijärjestuse tüüpide, geenide arvu ja vastastikuse paiknemise (sünteensuse, ahelduse, klastrilisuse) jms. järgi. Diploidse organismi gameedi haploidne (= monoploidne; vt. ploidsus) kromosoomistik sisaldab ühe genoomi (excl. gonosoomispetsiifilised lookused, millest esinevad kas X- või Y-liitelised, mitte mõlemad). Inimese Genoomi moodustavad 24 kromosoomi: 22 autosoomi ja 2 gonosoomi, X ja Y; selles sisaldub ca 3 miljardit nukleotiidipaari DNAd ning 22-25 tuhat geeni.
Genoomi mõistet kasut. ka rakuorganellide geneetilise struktuuri tähistamiseks (mitokondrigenoom, plastiidigenoom); need geneetilised struktuurid on tuumagenoomiga võrreldes väga väikesed (nt. inimese standardses mitokondrigenoomis 16569 np).
genoommutatsioon (genome mutation) -- indiviidi (raku) normaalse liigiomase kromosoomistiku arvuline muutus, kas kogu kromosoomikomplekti kordne muutus (euploidsus, polüploidsus) või üksikute kromosoomide arvu muutus (aneuploidsus).
genotüüp (genotype) -- 1) indiviidi (või raku) kogu geneetiline informatsioon, mis koostoimes keskkonnatingimustega määrab tema fenotüübi.
2) indiviidi (raku) geneetiliste lookuste alleelne koosseis. Diploidsetel organismidel eristatakse iga geeni suhtes homosügootset ja heterosügootset genotüüpi. Suguliselt sigivate diploidsete organismide genotüübi igas geenipaaris on üks alleel saadud isalt, teine emalt. Heterogameetse sugupoole genotüüp on suguliiteliste geenide suhtes hemisügootne.
genotüübiline muutlikkus (genotypic variability) -- indiviidide (rakkude) vaheliste erinevuste teke ja esinemine genotüüpides. Tekkepõhjuste järgi eristatakse mutatiivset ja kombinatiivset muutlikkust. Genotüübiline muutlikkus võib avalduda fenotüübilise muutlikkuse näol, kuid suur osa sellest on varjatud, st. fenotüüpides mitteavalduv (homo- ja heterosügootsusdominantsuse korral, sünonüümsed geenmutatsioonid, erinevused genoomide mittekodeerivates piirkondades).
gonosoom (gonosome) -- e. sugukromosoom -- kromosoom, mille olemasolu või arv on eri sugupooltel liigiomaselt erinev; gonosoomid määravad sügoodi geneetilise (kromosoomse) soo (vrd. autosoom). Enamikul loomadel ja osal taimedel on kromosoomistikus üks paar gonosoome, neid tähistatakse tähtedega (X ja Y või Z ja W). Sugupoolt, millel on ühesuguste (homoloogsete) gonosoomide paar, nim. homogameetseks (moodustab kromosoomistiku suhtes ühesuguseid gameete), neiks on nt. emasimetajad (XX) ja isaslinnud (ZZ); sugupoolt, millel on erinevad gonosoomid, nim. heterogameetseks (moodustab gonosoomide poolest erinevaid gameete), neiks on nt. isasimetajad (XY) ja emaslinnud (ZW). Enamasti on ainult heterogameetsele sugupoolele omane gonosoom (Y resp. W) märksa lühem ja sisaldab palju vähem geene kui mõlemale sugupoolele ühine kromosoom (X resp. Z). Isendite fenotüübilise soo determinatsiooni mehhanism on erinevates organismirühmades erinev, ka sama kromosoomse soomääramise tüübi korral. Nt. äädikakärbsel (Drosophila melanogaster) ja inimesel on mõlemal emassugu homogameetne (XX) ning isassugu heterogameetne (XY) ja sügoodi kromosoomne sugu määratakse viljastumisel spermitüübiga (X- või Y-gonosoomi sisaldavaga), ent fenotüübilise soo determinatsioon on põhimõtteliselt erinev. Äädikakärbse soo määrab autosoomikomplektide ja X-kromosoomide arvu suhe; Y-kromosoomide olemasolu või arv ei ole selle juures oluline. Inimese (nagu teistegi imetajate) puhul sõltub fenotüübilise soo areng (esmajoones gonaadisugu) Y-kromosoomist: kui esineb, areneb isasindiviid, kui puudub, siis emane. Ja sealjuures on Y-kromosoomi sugumäärav funktsioon tõenäoliselt seotud ainult ühe geeniga -- testiseid (e. seemnesarju) determineerivat faktorit (TDF) määrava geeniga (SRY).
haploidsus (haploidy) -- indiviidi (raku) kromosoomistiku poolkordsus, liigi haplofaasile omase (gameetse) kromosoomistiku olemasolu (arvsümbol n). Valdaval enamikul loomadel on haploidsus sama, mis monoploidsus (n = x, genoome üks), polüploidsetel liikidel võib haploidne kromosoomiarv mitmekordselt ületada monoploidset põhiarvu (n>x, genoome mitu). Suguliselt sigivatel organismidel vahelduvad elutsüklis haploidne ja diploidne faas. Vt. ploidsus.
haplotüüp (haplotype) -- 1) kitsamas tähenduses: mingi kromosoomi(lõigu) tihedalt aheldunud lookuste alleelne koosseis (nt vererühmade Rhesus-süsteemi alleelsed haplotüübid (CDe, cdE, CdE jne). 2) laiemas tähenduses: haploidne genotüüp, haploidse raku (gameedi, eose) või organismi paaritu genotüüp.
Hardy-Weinbergi seadus (Hardy-Weinberg law) -- e. populatsiooni geneetilise tasakaalu seadus e. populatsioonigeneetika põhiseadus. Suures isoleeritud panmiktilises populatsioonis, milles ei toimi mutagenees ega valik, säilivad alleeli- ja genotüübisagedused põlvkonniti muutumatutena, kusjuures genotüübisagedused on määratud alleelisagedustest ruutseose kaudu.
Kui lihtsaima mudelina vaadelda üht autosoomset dialleelset lookust, mille alleelide A ja a sagedused geenifondis on vastavalt p ja q, siis väljendab seost alleelisageduste ja genotüübisageduste vahel võrrand: [p(A) + q(a)]2 = p2(AA) + 2pq(Aa) + q2(aa) = 1. Populatsioon, kus kehtib see Hardy-Weinbergi võrrand, on geneetilise tasakaalu seisundis ja püsib selles, kuni mingi geneetilise dünaamika tegur (evolutsioonitegur) ei põhjusta muutusi.
Kui populatsioon ei ole tasakaaluseisus, siis nõutud tingimuste kehtestumisel läheb populatsioon iga autosoomse lookuse suhtes tasakaalu üheainsa põlvkonnavahetusega, kuid suguliiteliste (X-liiteliste) lookuste puhul saabub tasakaal mitme (taval. 10-13) põlvkonnavahetuse järel.
Tuleb rõhutada, et see seadus postuleerib kahte asjaolu: 1) alleeli- ja genotüübisageduste püsivust; 2) genotüübisageduste tasakaalulisust. Need näitajad võivad teatud juhtudel teineteisest sõltumatult muutuda. Näit. inbriiding muudab genotüübisagedusi mõjutamata alleelisagedusi ja viib populatsiooni geneetilisest tasakaalust välja. Kõik tegurid, mis muudavad alleelisagedusi (mutagenees, geenisiire, geenitriiv, valik), muudavad ka genotüübisagedusi, kuid võivad säilitada populatsiooni igas järjestikuses põlvkonnas tasakaaluseisu (st. Hardy-Weinbergi võrrand kehtib põlvest põlve, kuigi muutuvate sagedusväärtustega).
Hardy-Weinbergi seadus on populatsiooni geneetilise staatika ja dünaamika (mikroevolutsiooni) kirjeldamise ja prognoosimise alus.
hemisügootsus (hemizygosity) -- geenide (lookuste) esinemine genotüübis ühekordselt (mitte alleelipaaridena). Nii on heterogameetsel sugupoolel nii X- kui ka Y-liitelised geenid hemisügootses seisundis (st. puuduvad homo- ja heterosügootsed genotüübid ning alleelide dominantsus-retsessiivsus-suhted).
heterokromatiin (heterochromatin) -- vt. kromatiin.
heteroos (heterosis) -- e. hübriidjõud (hybrid vigour), nähtus, mis väljendub hübriidide (ristandite) suurenenud kasvus, kasvukiiruses, produktiivsuses, vastupidavuses jms. Heteroos ilmneb tavaliselt sama liigi inbriidsete liinide, vähemal määral tõugude või sortide ristamisel, aga mõnikord ka (sama perekonna) eri liikide hübriidimisel saadud indiviidide juures. Viimasel juhul ilmneb heteroos enamasti osaliselt: mõni omadus (taval. kasvukiirus, vastupidavus jms.) on võimendunud, kuid viljakus (sigimisvõime) on kas oluliselt langenud või kadunud; sellise heteroosi klassikaliseks näiteks on muul.
heterosügoot (heterozygote) -- heterosügootne (heterozygous) indiviid; vt. heterosügootsus.
heterosügootsus (heterozygosity) -- diploidse (või polüploidse) indiviidi genotüübi seisund, kus homoloogiliste kromosoomide samas lookuses (või mitmes vaatlusaluses lookuses) asuvad erinevad alleelid (nt. Aa). Vastandub homosügootsusele (homozygosity), mille puhul neis lookustes on identsed alleelid (nt. AA või aa). Tähistamaks heterosügootsust mitme erineva lookuse suhtes, kasutatakse termineid di-, tri-, polüheterosügootsus. Heterosügootse genotüübiga indiviidi nim. heterosügoodiks, homosügootset homosügoodiks.
holandriline (holandric) -- täielikult Y-liiteline geen (tunnus), mis pärandub ainult isalt poegadele ja avaldub üksnes isassool (isas-heterogameetsuse korral).
homoloogilised kromosoomid (homologous chromosomes) -- liigi kromosoomikomlekti mingi kromosoomi eksemplarid kas sama või eri indiviidide kromosoomistikus. Homoloogilised kromosoomid on kujult ja suuruselt sarnased ning sisaldavad enamasti samu geneetilisi lookusi samas järjestuses (erinevused tulenevad mõningate heterokromatiinsete lõikude polümorfismist ja harvadest kromosoommutatsioonidest - deletsioonide, inversioonide, translokatsioonide olemasolust). Homoloogiliste kromosoomide lookustes võivad olla samad või erinevad alleelid. Suguliselt sigivate organismide diploidne kromosoomistik koosneb homoloogiliste kromosoomide (homoloogide) paaridest, millest igaühes on üks kromosoom saadud isalt, teine emalt, ning meioosis need kromosoomid konjugeeruvad ja lahknevad eri gameetidesse või spooridesse.
homosügoot (homozygote) -- homosügootne (homozygous) indiviid; vt. heterosügootsus.
homosügootsus (homozygosity) -- vt. heterosügootsus.
inbriiddepressioon (inbreeding depression) -- indiviidide kohasuse ja elujõulisuse langus inbriidingu tagajärjel populatsioonis, kus normaalselt toimub panmiktiline ristumine. Inbriiddepressiooni põhjuseks peetakse suurenenud homosügootsuse astet paljude lookuste ebasoodsate retsessiivsete alleelide suhtes ja polügeensete regulatsioonisüsteemide tasakaalulisuse rikkumist. Erinevate inbriidsete liinide ristamisel ilmneb sageli heteroos.
inbriiding (inbreeding) -- igasugune ristumis- või ristamissüsteem, mille puhul paaruvate isendite sugulus on suurem, kui panmiktilises populatsioonis. Inbriidingu aste sõltub paaruvate isendite sugulusastmest; seda hinnatakse inbriidingukoefitsiendi (F) kaudu. Inbriidingu äärmusvorm on iseviljastumine (nt. taimede isetolmlemisel). Tavaliselt kasutatakse inbriidingu mõistet ristviljastuva liigi mittepanmiktilise sigimise puhul.
Inbriiding muudab populatsiooni genotüüpide suhtelisi sagedusi, kuid ei mõjuta selle geenifondis sisalduvate alleelide sagedusi. Seega saab rikutud genotüübisageduste tasakaaluline jaotus: suureneb homosügootide ja väheneb heterosügootide sagedus. Kui panmiktilises populatsioonis on autosoomse lookuse genotüüpide (AA, Aa, aa) tasakaalulised sagedused vastavalt p2, 2pq, q2, siis inbriidingu korral on need sagedused (p2 + pqF), (2pq - 2pqF), (q2 + pqF); (p ja q tähistavad alleelide A ja a sagedusi, F on inbriidingukoefitsient). Iseviljastumise korral fikseeruvad võrdlemisi kiiresti erinevad homosügootsed liinid, heterosügootsed genotüübid kaovad.
Sugulaste ristumise (sugulasabielude) korral üldiselt panmiktilises populatsioonis tähendavad viimati näidatud sagedused tõenäosusi, millega sünnivad sellistest paarumistest erinevate genotüüpidega järglased. Kuna retsessiivne fenotüüp (tunnus) avaldub üksnes homosügootse genotüübi puhul, siis põhjustab inbriiding eelkõige haruldaste retsessiivsete tunnuste (enamasti defektide, haiguste) tõenäosuse suurenemist sugulastepaari järglaste juures. See defektiriski suurenemine (võrreldes sagedusega panmiktilises populatsioonis) on väljendatav suhtega (q + pF)/q. Nagu nähtub, on see seda suurem, mida suurem on inbriidingukoefitsient (F) ja mida väiksem on retsessiivse alleeli sagedus (q). Ka sel juhul, kui inbriidsete indiviidide juures ei ilmne konkreetseid tunnusepuudeid (resp. haigusi), on nad tavaliselt väiksema kohasusega; seda nim. inbriiddepressiooniks.
inbriidingukoefitsient (inbreeding coefficient) -- inbriidingu astme mõõt populatsioonis või sugulasisendite ristumisel (resp. ristamisel) saadud järglaste juures; tähistatakse tavaliselt sümboliga F. Otseselt saab inbriidingukoefitsienti määrata sugulasisendite ristumisel (resp. sugulasabielust) sündinud järglaste jaoks: see on tõenäosus, et isend on homosügootne päritolu poolest identsete (so. vanemate ühiselt eellaselt pärit) alleelide suhtes. Suures panmiktilises populatsioonis F = 0. Sugulasristamisest (-abielust) sündinud indiviidi inbriidingukoefitsient arvutatakse järgmise valemi kohaselt:
F = (1/2)n-1, kus n on rajaetappide (põlvkondade) arv vanemate ühise eellase juurde üle kummagi vanema; kui vanematel on rohkem kui üks ühine eellane, siis tuleb need väärtused eraldi arvutada ja summeerida. Populatsioni keskmine inbriidingukoefitsient arvutatakse kõigist sugulasristumistest sündivate järglaste suhtosa ja nende keskmise inbriidingukoefitsiendi järgi.
Inbriidingukoetsiendi väärtusi erineva sugulusastmega vanemate järglastele:
Vanemate sugulus F
Iseviljastumine 1/2 = 0,5
Isa-tütar/Ema-poeg 1/4 = 0,25
Õved (õde-vend) 1/4 = 0,25
Poolõved 1/8 = 0,125
Onu - õe/vennatütar 1/8 = 0,125
Nõod (õvede lapsed) 1/16 = 0,0625
Poolnõod 1/32 = 0,03125
Teise astme nõod 1/64 = 0,0156
Vanemate kaugema suguluse puhul pole järglaste inbriidingukoefitsiendil olulist tähendust.
intermediaarsus (intermediary) -- intermediaarne alleelitoime, tunnuse vahepealne avaldumine heterosügootse genotüübi puhul võrreldes mõlemal homosügoodil esineva tunnusega. Osa geneetikuid nimetab seda nähtust mittetäielikuks domineerimiseks.
Nähtuse olemus seisneb geeni (alleeli) doosiefektis.
juhuslik sündmus (random event) -- sündmus, mis antud tingimustes ei ole ühetähenduslikult determineeritud, st. võib toimuda teatud tõenäosusega P (0<P<1). Heterosügootide järglaskonnas võimalikud genotüübid on iga üksikisendi jaoks juhuslikud (tõenäosuslikud) sündmused.
karüogramm (karyogram) -- indiviidi karüotüübi uurimiseks kasutatav kromosoomistiku süstematiseeritud fotokujutis ühe raku metafaasikromosoomidest, kus kromosoomipaarid on reastatud ja rühmitatud suuruse, kuju ja vöödistusmustri järgi. Inimese autosoome tähistatakse suuruse alanevas järjekorras numbritega 1...22 ja jaotatakse suuruse ning kuju alusel 7sse rühma A...G (A: suured metatsentrikud, B: suured submetatsentrikud, C: keskmised submetatsentrikud, D: suured akrotsentrikud, E: väikesed submetatsentrikud, F: väikesed metatsentrikud, G: väikesed akrotsentrikud), gonosoome tähistatakse tähtedega X ja Y.
karüotüübianomaalia (karyotype abnormality) -- indiviidi (raku) ebanormaalne karüotüüp; kromosoomide arvu, suuruse ja kuju kõrvalekalded liigiomastest varieeruvuspiiridest. Karüotüübianomaaliad on tingitud kromosoommutatsioonidest (deletsioonid, duplikatsioonid, translokatsioonid, inversioonid) ja genoommutatsioonidest (aneuploidsus nagu monosoomia, trisoomia jne., polüploidsus); nad põhjustavad nn. kromosoomhaigusi (nt. Down'i, Klinefelter'i ja Turner'i, kassikisa jm. sündroomid inimesel).
karüotüüp (karyotype) -- indiviidi kromosoomistiku tunnustekogum, mida iseloomustab kromosoomide arv, suurus, (tsentromeeri asukohast olenev) kuju ja värvumismuster (vöödistus). Kui organismis on erineva kromosoomiarvu või -struktuuriga rakke, siis on indiviidil mosaiikkarüotüüp. Karüotüüpi uuritakse ja iseloomustatakse tavaliselt mitoosi metafaasi (või prometafaasi) kromosoomistikul karüogrammi koostamise abil.
kloon (clone) -- organismide, rakkude ja DNA-molekulide geneetiliselt identne kogum, mis on tekkinud ühest eellasest replikatsiooni ja suguta (rekombinatsioonivaba) paljunemise teel. 1) organismikloon: isendi suguta sigimisel (apogaamia puhul, vegetatiivsel paljunemisel ja paljundamisel, somaatilise rakutuuma siirdamisel) tekkinud isendite kogum, kes on geneetiliselt identsed omavahel ja vanemorganismiga. Loomade ja inimese ühest viljastunud munarakust arenevad ühemuna- e. monosügootsed mitmikud moodustavad samuti klooni (tekkinud embrüo vagetatiivse paljunemise e. polüembrüoonia teel). 2) rakukloon: hulkrakse organismi geneetiliselt või epigeneetiliselt erilaadse raku mitootilisel jagunemisel (in vivo või in vitro) tekkinud rakukogum. 3) DNA-kloon: bakteri plasmiidi või pärmseene 'kunstlikku kromosoomi' siirdatud inimese või muude eukarüootide DNA-fragmentide (geenide) kogum nende organismide kloonis.
kloonimine (cloning) -- organismi-, raku- või DNA-kloonide tekitamine uurimis- ja aretustöö ning tootmise eesmärgil.
1. Organismikloonimine. Taimede kloonimise põhimeetod seisneb paljundamises vegetatiivorganite (mugulate, sibulate, risoomide, varte, lehtede, pungade jms.) abil. Kõrgemate taimede kloonimiseks (eriti viirusvabade liinide saamiseks) kasutatakse nüüdisajal ka meristeemmeetodit: taimi kasvatatakse üles kasvukuhiku algkoe (meristeemi) tükikestest koekultuuris. Imetajate kloonimiseks on kaks võimalust: (1) embrüokloonimine -- varajane embrüo jagatakse üksikuteks blastomeerideks või suuremateks osadeks, neid kasvatatakse in vitro ja ja saadud tütarembrüod siirdatakse siirdatakse hormonaalselt töödeldud retsipientlooma emakasse. Seda meetodit rakendatakse koduloomade kloonimisel alates 1970ndatest aastatest. (2) tuumkloonimine -- täiskasvanud looma mingi keharaku tuum siirdatakse munarakku, millest on oma tuum eemaldatud. Sel juhul saadakse tuumadoonoriga geneetiliselt (enam-vähem ?) identsed järglased. Esimene edukas imetaja tuumkloonimine sooritati Edinburghis Roslini Instituudis I. Wilmuti juhtimisel 1996. a. (sündis kloonitud lammas Dolly). Samas instituudis teostati inimese geeni kandva siirdgeense lamba kloonimine 1997. a. (sündis kloonlammas Polly). 2. Rakukloonimine. Hulkraksete loomade rakkude kloonimiseks külvatakse hõredaid rakususpensioone pooltahkesse söötmesse või plastikplaadi lohkudesse või eraldatakse üksikud rakud mikropipetiga ja kultiveeritakse eraldi paljundussöötmes. Nii saadakse nt. monokloonseid antikehi produtseerivaid hübridoomirakkude kloone. 3. DNA-kloonimine. Inimese (või muude organismide) suvalisi või kindlaid geene sisaldavaid DNA-fragmente kloonitakse bakterikloonide plasmiidides või pärmseenerakkude kunstlikes kromosoomides. Sel meetodil on loodud inimese genoomi mitmesugused pangad.
kodominantsus (codominance) -- tunnuste (alleelide) võrdväärne ja teineteisest sõltumatu avaldumine heterosügootse genotüübi puhul; mõlemast alleelist määratud tunnusevariandi üheaegne avaldumine heterosügootse indiviidi (raku) fenotüübis. Alleelide kodominantne avaldumine on tavaline molekulaarsete tunnuste, sh. rakupinna antigeenide (nt. vererühmade) puhul.
kohasus (fitness) -- e. valikuväärtus (selective value) e. adaptiivväärtus (adaptive value), mingi genotüübiga isendite suhteline sigimisedukus võrreldes antud populatsiooni teiste genotüüpidega. Kohasust tähistatakse populatsioonigeneetikas sümboliga w. Iga genotüübi kohasuse määramisel hinnatakse selle sigimisedukust kõige võimekama suhtes, kusjuures kriteeriumiks on sigimisvõimeliste järglaste arv. Maksimaalse sigimisedukuse korral on kohasuse väärtus 1 (w = 1), geneetilise letaalsuse korral (puuduvad sigivõimelised järglased) on kohasus null (w = 0). Kohasus sõltub eluvõimest ja viljakusest. Kohasuse määranguid kasutatakse loodusliku valiku intensiivsuse hindamisel eri genotüüpide suhtes (tavaliselt üksikute lookuste kaupa) populatsiooni geneetilise dünaamika e. mikroevolutsiooni uurimisel. Kohasuse vastandväärtuseks on selektsioonikoefitsient (s), millega hinnatakse mingi genotüübi suhtelise eliminatsiooni intensiivsust. Kohasus ja selektsioonikoefitsient on täiendsuurused (w + s = 1; w = 1 - s; s = 1 - w), nii et populatsioonigeneetilistes võrrandites võib kasutada üht teise asemel.
kombinatiivne muutlikkus (segregational variation) -- sugulise sigimisega kaasnev geneetiline muutlikkus; tuleneb kromosoomide ja geenipaaride meiootilisest rekombinatsioonist ning gameetide juhuslikust paardumisest viljastumisel. Kombinatiivset muutlikkust eukarüootidel kirjeldavad Mendeli ja Morgani seadused.
kromatiid (chromatid) -- üks kahest mitoosi- ja meioosiaegse kromosoomi pikipoolmest, õdekromatiidist e. tütarkromatiidist. Kromosoomi kahekromatiidilisus tekib ühekromatiidilise kromosoomi reduplikatsiooni tulemusena; see põhineb interfaasi S-perioodis toimunud DNA replikatsioonil. Nad eralduvad mitoosi anafaasis või meioosi II anafaasis ja liiguvad iseseisvate (ühekromatiidiliste) kromosoomidena raku eri poolustele.
kromatiin (chromatin) -- eukarüootsetekromosoomide koostisaine, nukleoproteiidne kompleks. Seda mõistet kasutatakse tavaliselt interfaasse tuuma kromosoomse materjali kohta, kus tervikkromosoomid ei ole tuvastatavad. Kromatiin ilmneb seal aluseliste kromatiinivärvidega värvuvate terakeste, känkude (kehakeste) ja fibrillidena, mis moodustab tihedamaid masse tuumakatte all (st. tuuma perifeerias). Kromatiin koosneb peamiselt DNAst ja sellega seotud valkudest: DNAga massivõrdses hulgas aluselistest kromatiinivalkudest histoonidest ja varieeruvas hulgas ning muutliku koosseisuga happelistest valkudest (e. hertoonidest); peale nende on kromatiinis RNAd (seal toimub selle süntees). Eristatakse eu - ja heterokromatiini. Eukromatiin värvub interfaasituumas ja mitoosi- ning meioosikromosoomides nõrgalt, selles on DNA vähemal määral kokku pakitud ja histoonide kõrval rohkem happelisi valke; eukromatiinseis kromosoomiosades asuvad aktiivsed geenid. Heterokromatiinis on DNA tihedamalt kokku pakitud ning selles on vähe happelisi valke; see moodustab peamise osa interfaasituumas tugevalt värvuvast kromatiinist; selles asuvad ka tandeemselt kõrgkorduvad satelliit-DNA järjestused. Vt. ka sugukromatiin.
kromomeer (chromomere) -- üks paljudest kromatiidi tihedamatest, piki kromatiidi helmesjalt paiknevatest paksenditest, mis on vaadeldavad meioosi ja (mõnevõrra ebaselgemalt) mitoosi profaasi kromosoomides. Kromomeeride ja nende vahemike (interkromomeeride) suurus on eri kromosoomiregioonides erinev. Kromomeerides on DNA tihedamalt kokku pakitud kui interkromomeerides.
kromoneem (chromonema) -- peeneim valgusmikroskoobis nähtav kromosoominiit interfaasi tuumas (läbimõõt u. 300 nm).
kromosoom (chromosome) -- 1) eukarüootse organismi (tuuma)genoomi struktuurselt individuaalne element, niitjas nukleoproteiidne moodustis rakutuumas, milles asuvad lineaarses järgnevuses ja kindla paiknevusega geenid ning mitmesugused mittegeensed nuleotiidijärjestused (lookused). Kromosoom sisaldab ühe DNA molekuli, sellega massivõrdses koguses aluselisi valke -- histoone, varieeruvas hulgas mittehistoonseid (happelisi) valke ja vähesel hulgal RNAd; seda kromosoomi koostisainete kompleksi nim. kromatiiniks. Kromosoomid läbivad mitoosi ja meioosi eel ning ajal keerukaid reproduktsiooni, kondensatsiooni-dekondensatsiooni ja lahknemise protsesse. Mitoosi (ja meioosi) pro- ja metafaasis koosneb kromosoom kahest kromatiidist (õdekromosoomist), mille ühendus katkeb viimasena tsentromeeri kohalt. Tsentromeer (e. primaarsoonis) jagab kromosoomi tavaliselt kaheks osaks (õlaks); mõnes kromosoomis esineb sekundaarsoonis, mis eraldab väikese (keraja) tipuosa, nn. satelliidi. Meioosi (ja mõnevõrra ebaselgemalt mitoosi) profaasis ilmnevad piki kromosoomi (kromatiidi) tihedamad helmesjad paksendid -- kromomeerid. Mikroskoopiliselt vaadeldavate kromosoomide erivormideks on lambihari- ja polüteensed kromosoomid.
2) sageli nim. kromosoomiks ka prokarüootide ja rakuorganellide (mitokondrite, kloroplastide) genoomi (e. genofoori), mille moodustab üks rõngasjas DNA-molekul ning millel ei ole kromatiinset püsiühendust valkudega (histoonid puuduvad).
kromosoomikomplekt (chromosome set) -- liigiomaste kromosoomide täielik ühekordne komplekt, milles iga kromosoom moodustab vähemalt ühe aheldusrühma (aheldus). Kromosoomikomplektis sisalduvad geenid ja muud lookused (järjestusmarkerid) moodustavad genoomi. Inimese kromosoomikomplekti kuulub 24 kromosoomi: 22 autosoomi ja 2 gonosoomi (X ja Y). Kromosoomikomplekti autosoomid ja üks gonosoom (kui organismil gonosoomid esinevad) on minimaalne (monoploidne) kromosoomistik, mis iga normaalse gameedi kaudu järglastele pärandatakse; seda kromosoomiarvu nim. liigi kromosoomide põhiarvuks.
kromosoomistik (chromosome complement -- kromosoomide kogum indiviidi somaatilise või generatiivse raku tuumas; võib sisaldada erineva arvu liigiomaseid kromosoomikomplekte - kas ühe (monoploidsus, haploidsus), kaks (diploidsus) või mitu (polüploidsus) või olla hüpo- ja hüperploidne (aneuploidsus). Indiviidi K-u tunnustekogumit, mida kirjeldatakse kromosoomide arvu, suuruse, kuju ja struktuuri kaudu, nim. karüotüübiks.
kromosoomiõlg (chromosome arm) -- (e. -haar) tsentromeeriga eraldatud kromosoomiosa; tavaliselt on kromosoomil kaks õlga (telomeersel kromosoomil üks), enamasti on üks õlg pikem (tähistatakse q) kui teine (p); nende pikkussuhte alusel eristatakse inimesel meta-, submeta- ja akrotsentrilisi kromosoome (viimastel on lühikese õla tipus satelliit, excl. Y-kromosoomil).
kromosoommutatsioon (chromosome mutation) -- indiviidi karüotüübi tsütoloogilise uurimisega tuvastatav kromosoomi struktuuri (pikkuse, kuju, vöötmustri) muutus, mis võib olla kas deletsioon, translokatsioon, duplikatsioon või inversioon (nende puhul eristatakse veel makro- ja mikrovariante).
kunstlik valik (artificial selection) -- eksperimentaatori või aretaja rakendatav valikuline toime (eliminatsioon, reproduktsiooni piiramine või soodustamine) bioloogilistele objektidele (rakkudele, indiviididele) eesmärgiga saada teatud vajadustele või soovidele ja rakendatud tingimustele vastavaid vorme (liine, tüvesid, sorte, tõuge). Suguliselt sigivate organismide (koduloomade, kultuurtaimede) puhul seisneb kunstlik valik peamiselt soovitud fenotüüpidega (või genotüüpidega) isendite kontrollitud ristamises ning järglaste valikus geneetiliste omaduste järgi (mittesoovitavate omadustega isendid kõrvaldatakse paljundusest).
lahknemisseadus (law of segregation) -- Mendeli II seadus (Mendel's second law).
laionisatsioon (lyonization) -- vt. geenidoosi kompenstasioon.
lambiharikromosoom (lampbrush chromosome) -- erivorm hiidkromosoome paljude (nii selgrootute kui ka selgroogsete) loomade ovotsüütitide ja Drosophila spermatototsüütide meioosi profaasi keskosas (diploteenis). Lambiharikromosoomid avastati esmalt sabakonnaliste ovotsüütides, kus nad ongi kõige suuremad. Neid kromosoome iseloomustavad kromatiidide paljudest kromomeeridest lateraalselt välja ulatuvad varieeruva pikkusega ja ajas muutuva paiknevusega lingud. Iga ling on tõenäoliselt üks transkriptsiooniüksus -- geen -- , millelt toimub aktiivne transkriptsioon; need lingude hargnemiskohad on võrreldavad polüteensete kromosoomide puhvidega. [Termini päritolu: 19. sajandi lõpu tsütoloogidele meenutasid need kromosoomid lambiharja; lambihari oli petrooleumilambi klaasi puhastamise vahend; tänapäeval tuntud vahenditest on sellele kõige sarnasem pudelihari].
looduslik valik (natural selection) -- looduses toimuv protsess, mis seisneb kohasemate geneetiliste variantide (organellide, rakkude, indiviidide, deemide) edukamas üleelamises ja paljunemises ja vähemkohasete (puuetega) variantide eliminatsioonis (suuremas suremuses või piiratud reproduktsioonis) antud ökoloogilistes tingimustes. Loodusliku valiku mõiste võttis kasutusele Ch. Darwin, seletamaks liikide evolutsiooni kohastumuslikult suunatud iseloomu; selle aluseks on looduses toimiv olelusvõitlus. Darvinliku loodusliku valiku toime järgi populatsiooni geneetilisele struktuurile eristatakse mitmeid valikuvorme, tavaliselt kolm põhitüüpi: stabiliseeriv, suunav ja lõhestav valik. Peale selle on valikutegurite ja mehhanismide alusel eristatavad veel suguline valik, sagedusest sõltuv valik ja rühmavalik. Valiku toime populatsiooni geneetilisele struktuurile sõltub erinevate genotüüpide kohasusest, mida populatsiooni geneetilise dünaamika modelleerimisel hinnatakse tavaliselt üksikute lookuste kaupa. Valiku mõju alternatiivsete tunnuste sagedusele (nende eliminatsioonile ja fikseerimisele või tasakaalulisele polümorfismile) oleneb tunnuste geneetilise määratuse tüübist (dominantsed või retsessiivsed tunnused) ja sellest, kas valik eristab heterosügoote ja homosügoote (nt. valik heterosügootide vastu või kasuks).
lookus (locus, mitm. loci) -- algne tähendus: geeni asukoht kromosoomigeneetilisel kaardil. Klassikalises geneetikas, eriti populatsioonigeneetikas kasutatakse termineid lookus ja geen tihti sünonüümidena. Viimasel ajal käibib laiem tähendus: mingil viisil (hrl. alleelse muutlikkuse järgi, vt. alleel) iseloomustatav (identifitseeritav) kromosoomi- või DNA-molekuli lõik (-sait), milles paikneb kindel geen (geenilookus) või mis tahes muu eristatav nukleotiidijärjestus (marker).
meioos (meiosis) -- organismide sugulise sigimise tsüklis toimuv kahekordne rakujagunemine, mille tulemusel diploidnekromosoomistik redutseerub haploidseks (või tetraploidne diploidseks); meioosis toimub vanematelt päritud kromosoomide (ja geenide) ümberkombineerumine uuteks haploidseteks kombinatsioonideks e. haplotüüpideks (so. geneetiline rekombinatsioon). Meioosi kahele jagunemisele eelneb üks S-perioodiga interfaas; kummaski jagunemises eristatakse (nagu mitoosiski) pro-, meta-, ana- ja telofaasi. Meioosi geneetilised iseärasused tingib homoloogiliste kromosoomide konjugatsioon (süünaps) esimese jagunemise profaasis, millest tuleneb geneetilise ristsiirde võimalus ja homoloogide lahknemine esimeses anafaasis. Loomadel toimub meioos gametogeneesis, taimedel sporogeneesis; enamikul seentel on meioos sügoodi esimene jagunemine. Meioosi ja viljastumise vaheldumine jagab suguliselt sigivate organismide elutsükli kahte faasi: haplo- ja diplofaasi (taimedel gameto- ja sporofüüdi põlvkonna vaheldumise kujul). Vrd. mitoos.
Mendeli seadused (Mendel's laws) - eukarüootide tuumakromosoomide (aheldumata) geenide pärandumise ja kombineerumise peamised seaduspärasused. Avastatud esmakordselt G. Mendeli poolt (1865) ja üleavastatud H. de Vries'i, C. Correns'i ja E. v. Tschermak'i poolt (1900). Enamasti defineeritakse kolm M. s-t: ühetaolisuse, lahknemise ja sõltumatu lahknemise seadus.
Mendeli I seadus (Mendel's first law) -- e. ühetaolisuse seadus (law of uniformity). Erinevate homosügootsete (e. pärivuspuhaste) isendite ristamisel on esimese põlvkonna järglased (F1) kõik ühetaolised heterosügoodid (sõltumata ristamise suunast).
Seadust võib väljendada genotüübivalemite abil: AA x aa --> Aa või aaBB x AAbb --> AaBb. Järglaste ühtlik fenotüüp (tunnuselaad) sõltub alleelide vastasmõjust. Üksiktunnus võib olla ühe vanema tunnuse sarnane (dominantsus, esineb enamiku alternatiivtunnuste puhul) või vanemate tunnuste vahepealne (intermediaarsus) või mõlema vanema tunnuse ühendus (kodominantsus). Polühübriidsest ristamisest saadud järglased sarnanevad rohkem sellele vanemale, kummal on rohkem dominantseid tunnuseid, kuid nad võivad olla ka kõigi tunnuste suhtes mõlema vanema vahepealsed.
Mendeli II seadus (Mendel's second law) -- e. lahknemisseadus (law of segregation). Heterosügootide (hübriidide) järglaskonnas toimub geneetiline lahknemine, nii et kindlates sagedussuhetes tekivad nii homosügootsed kui ka heterosügootsed isendid.
Algselt on seadus formuleeritud monohübriidide ristamisest saadud teise hübriidpõlvkonna lahknemise seaduspärasusena:
F1: Aa x Aa --> F2: 1/4AA + 2/4Aa + 1/4aa. Saadava kolme genotüübi sageduste 1:2:1-suhe tuleneb alleelide lahknemisest erinevatesse gameetidesse meioosis ja nende vabast paardumisest viljastumisel, mis on väljendatav järgmise avaldisega: Aa x Aa --> (1/2A + 1/2a) x (1/2A + 1/2a) --> 1/4AA + 2/4Aa + 1/4aa. Kirjeldatud genotüübiline 1:2:1-lahknemine võib avalduda erinevate fenotüübiliste lahknemistena (kas 3:1- või 1:2:1-suhteis), olenevalt alleelide interaktsioonist: 3/4 dominantseid + 1/4 retsessiivseid või 1/4 tunnusega 1 + 2/4 vahepealseid + 1/4 tunnusega 2. Lahknemisseadus toimib aga ka siis, kui ainult üks vanem on heterosügootne, ja ka siis, kui vanemad on erinevad heterosügoodid: Aa x aa --> 1/2Aa + 1/2aa või (trialleelsuse korral) A1a x A2a --> 1/4A1A2 + 1/4A1a + 1/4A2a + 1/4aa. Lahknemisseadus on statistiline seaduspärasus, mistõttu teoreetilistele sagedustele lähedased suhted on vaadeldavad järglaskonna piisavalt suure arvukuse korral (eri vanempaaride järglaste summeerimisel). Üksikjärglaste genotüüpide ja fenotüüpide jaoks tähendavad nad tõenäosusi.
Mendeli III seadus (Mendel's third law) -- sõltumatusseadus (law of independence) e. sõltumatu lahknemise seadus (law of independent assortment) e. vaba kombineerumise seadus (law of free combination). Polüheterosügootide erinevad alleelipaarid lahknevad ja kombineeruvad üksteisest sõltumatult.
Selle seaduse kohaselt moodustab polüheterosügoot (e. polühübriid) võrdse sagedusega 2n erineva haplotüübiga gameeti (kus n on heterosügootsete geenipaaride arv e. hübriidsusaste). Nii moodustab dihübriid 4 tüüpi gameete: AaBb --> 1/4AB + 1/4Ab + 1/4aB + 1/4ab; trihübriid 8 tüüpi: AaBbDd --> 1/8ABD + 1/8ABd + 1/8AbD + 1/8Abd + 1/8aBD + 1/8aBd + 1/8abD + 1/8abd. Erinevate isas- ja emasgameetide vabal viljastumispaardumisel võib tekkida 3n erineva genotüübiga sügooti kindlate sagedustega. Polühübriidide järglaskonnas ilmnevat geno- ja fenotüübilistlahknemist saab mõista üksikute alleelipaaride (ja tunnuste) suhtes tekkivate lahknemisridade vaba kombineerumisena: AaBb x AaBb --> (1/4AA + 2/4Aa + 1/4aa) x (1/4BB + 2/4Bb + 1/4bb) --> 1/16AABB + 2/16AABb + 1/16AAbb + 2/16AaBB + 4/16AaBb + 2/16Aabb + 1/16aaBB + 2/16aaBb + 1/16aabb. Kui mõlemas heterosügootses geenipaaris esineb domineerimine, siis saame dihübriidse fenotüübilise lahknemise leida järgmiselt:
(3/4Dom1 + 1/4rets1) x (3/4Dom2 + 1/4rets2) --> 9/16Dom1,Dom2 + 3/16Dom1,rets2 + 3/16rets1,Dom2 + 1/16rets1,rets2, kus Dom ja rets tähistavad kummaski paaris vastavalt dominantset ja retsessiivset tunnust.
Polüheterosügootide järglaste võimalike genotüüpide (ja fenotüüpide) tõenäosusi saab arvutada üksikute alleelipaaride (või tunnuste) tõenäosuste korrutamise teel: olgu vanemate genotüübid AabbCcDdEe x AaBbCcDDEe; genotüübi AAbbCcDDee tõenäosus oodataval järglasel on 1/4 x 1/2 x 1/2 x 1/2 x 1/4 = 1/128.
Sõltumatu lahknemise seadus ei kehti aheldunud lookuste korral (vt. geneetiline aheldus).
mendelism (mendelism)-- teadusliku geneetika algne nimetus; geneetika alusteooria, mis väidab, et pärilikkus põhineb püsivatel ja sugurakkude moodustumise ning viljastumise käigus lahknevatel ja kombineeruvatel pärivusteguritel, geenidel, mis määravad kindlate tunnuste kujunemise. (Vt. Mendeli seadused).
M-i põhikontseptsioonideks on pärilikkuse geeni-, kromosoomi- ja mutatsiooniteooria. M. on nimetatud Gregor Mendeli (1822-1884) järgi, kes oli Brnos (Tshehhimaal) augustiinlaste kloostri munk, kooliõpetaja ja loodusteadlane, pärilikkuse geeniteooria rajaja (põhitöö "Versuche über Pflanzen-Hybriden" ilmus 1866, Brünni Looduseuurijate Seltsi toimetiste 1865. a. köites).
mitoos (mitosis) -- eukarüootsete rakkude jagunemise põhiviis, mille puhul emarakust tekib kaks samasuguse kromosoomistikuga tütarrakku. Mitootiliselt jagunevad rakud läbivad rakutsükleid (e. mitoositsükleid), milles järgnevad interfaas (koosneb omakorda G1-, S- ja G2-perioodist) ning mitoosifaasid (pro-, meta-, ana- ja telofaas). S-perioodis replitseerub DNA ja kahestuvad kromosoomid -- tekivad kromatiidid, mis anafaasis lahknevad tütarkromosoomidena. Mitootiliselt võivad jaguneda igasuguse kromosoomiarvuga rakud. Mitoosil põhineb organismide vegetatiivne sigimine, hulkrakse organismi kasv, areng ja kudede uuendumine. Vrd. meioos.
mittetäielik dominantsus (incomplete dominance) -- tunnuse avaldumine heterosügootse genotüübi puhul erinevalt mõlemast homosügoodist, kuid kalduvusega ühe (dominantse) homosügoodi poole. Osa geneetikuid nimetab mittetäielikuks dominantsuseks kõiki juhte, kus heterosügoodi tunnus on homosügootide tunnuste vahepealne (vt intermediaarsus).
modifikatsioon (modification) -- mingist keskkonnatingimusest põhjustatud mittepärilik muutus indiviidi fenotüübis. Vt. modifikatsiooniline muutlikkus.
modifikatsiooniline muutlikkus (modicational variability) -- modifikatsioonide teke ja esinemine antud genotüübiga isendite
fenotüüpides. Modifikatsiooniline muutlikkus tuleneb genotüüpide erinevast avaldumisest erinevates keskkonnatingimustes. Modifikatsioonilise muutlikkuse kvalitatiivne iseloom ja kvantitatiivne ulatus on määratud päriliku reaktsiooninormiga. Erineva genotüübiga isendite sama tunnuse ja ühe isendi erinevate tunnuste modifikatsiooniline muutlikkus on erineva reaktsiooninormiga; seda tähistatakse tihti mõistega fleksibiilsus. Tunnuste modifikatsiooniline muutlikkus oleneb ka toimiva keskkonnateguri ajast indiviidi ontogeneesis, st. eri tunnustel esinevad erinevad kriitilised (tundlikud) perioodid.
monoploidsus (monoploidy) -- liigiomase kromosoomikomplekti (kõik autosoomid ja üks gonosoom) ühekordsus indiviidi (raku) kromosoomistikus. Monoploidset kromosoomiarvu nim. liigi kromosoomide põhiarvuks (sümbol: x), mis on väikseim haploidne arv polüploidseerias. Diploidsete organismide (valdava enamiku loomade ja paljude taimede) puhul on monoploidsus sama, mis haploidsus. Monoploidne kromosoomistik sisaldab ühe genoomi (excl gonosoomide mittehomoloogsed osad, millest kuuluvad sellesse ainult X- või Y-liitelised lookused, mitte mõlemad).
Morgani seadus (Morgan's law) -- aheldusseadus (law of linkage) seaduspärasus, mis tuleneb geenide (ja muude geneetiliste markerite) jadalisest paiknemisest kromosoomides. Seda seadust defineeritakse alternatiivsete reeglitena:
- aheldusreegel: ühes kromosoomis lähestikku paiknevad geenid on lineaarses ahelduses ja päranduvad enamasti koos, sealjuures seda sagedamini, mida väiksem on nende vahemaa piki kromosoomi;
- ristsiirdereegel: aheldunud geenid rekombineeruvad meiootilise ristsiirde kaudu, kusjuures nende ümberkombineerumise sagedus on seda suurem, mida suurem on geenide vahemaa piki kromosoomi.
mutabiilsus (mutability) -- replitseeruvate geneetiliste struktuuride (geenide, genoomide) omadus muutuda, muteeruda (mutatsioon). Mutabiilsusest tulenevad esmased geneetilised muutused biosüsteemides, millest johtub igasugune pärilik mitmekesisus ja evolutsiooni võimalus. Mutabiilsuse põhjuseks on geneetiliste elementide replikatsiooni, rekombinatsiooni ja lahknemise vead. Eri organismide ja sama genoomi eri osade (lookuste) mutabiilsus on tihti erinev.
mutageen (mutagen) -- mistahes füüsikaline või keemiline tegur, mis suurendab mutatsioonide sagedust üle spontaanse mutabiilsuse taseme. Mutageenidega mõjustamisel saadud mutatsioone nim. indutseeritud mutatsioonideks, vastandatuna normaaltingimustes tekkivatele spontaansetele mutatsioonidele. Mutageenid võivad tõsta mutatsioonisagedust kuni sadu ja tuhandeid kordi. Tuntumad mutageenid on radioaktiivne, röntgen- ja UV-kiirgus, lämmastikaluste analoogid ja alküleerivad ühendid, mis võivad indutseerida nii geen- kui ka kromosoommutatsioone. Indutseeritud genoommutatsioonide põhjuseks on sageli käävimürgid, nt. kolhitsiin.
mutagenees (mutagenesis) -- mutatsioonide tekke protsess, spontaanne või indutseeritud. Mutageneesi intensiivsus oleneb organismi geneetilistest omadustest (nt. DNA reparaaside aktiivsusest) või välistest teguritest (mutageenidest). Vt. ka mutabiilsus.
mutant (mutant) -- mutatsioonist põhjustatud fenotüübimuutusega indiviid; indiviid, kes on pärilikult erinev looduslikus populatsioonis levinud tavavormidest (nn. metstüüpi isendeist).
mutatiivne muutlikkus (mutative variability) -- e. mutatsiooniline muutlikkus, geneetilise muutlikkuse osa, mis on tingitud mutageneesist populatsioonis.
mutatsioon (mutation) -- indiviidi (raku, organelli, viiruse) geneetilise struktuuri (geeni, kromosoomi, genoomi) muutus, mis pärandub tütarrakkudele või ka järglaspõlvkonna indiviididele (va. dominantne letaalmutatsioon ja somaatiline mutatsioon mitte-vegetatiivselt sigivatel organismidel) ja põhjustab sageli muutunud fenotüübiga raku/indiviidi (mutandi) tekke. Suur osa mutatsioone on organismidele kahjulikud (sh. letaalsed), kuid on ka palju neutraalseid ja isegi kasulikke mutatsioone (mutatsiooni adaptiivväärtus võib oleneda organismi elukeskkonnast). Muutunud geneetilise struktuuri järgi eristatakse geenmutatsioone (DNA nukleotiidijärjestuse muutus üksiku geeni piires), kromosoommutatsioone (mikroskoopiliselt identifitseeritavad muutused kromosoomide struktuuris -- translokatsioonid, deletsioonid, inversioonid) ja genoommutatsioone(kromosoomide arvu muutused liigiomases kromosoomistikus). Tekkekoha järgi organismi kudedes eristatakse somaatilisi ja generatiivseid mutatsioone; tekkepõhjuste järgi spontaanseid ja indutseeritud mutatsioone. Spontaansete geenmutatsioonide sagedus on väike, tavaliselt vahemikus 10-6 ...10-5 geeni kohta põlvkonnas; spontaansete genoommutatsioonide tekkesagedus on kõige suurem, ca 10-3 kromosoomi kohta põlvkonnas.
muutlikkus (variability, variation) -- erinevuste teke või olemasolu biosüsteemide (organismide, rakkude, organellide, viiruste, populatsioonide, liikide) vahel. Muutlikkus on mitmetasandiline ja erineva olemusega. Nii võib eristada individualset ja rühmaviisilist (sh. taksonoomilist) muutlikkust. Individuaalses muutlikkuses eristatakse enamasti genotüübilist ja fenotüübilist, geneetilist ja mittegeneetilist (modifikatsioonilist), alternatiivset (kvalitatiivset, diskreetset) ja pidevat (kvantitatiivset), normaalset ja patoloogilist muutlikkust.
nullisoomsus (nullisomy) -- aneuploidsuse vorm, mille puhul indiviidi (raku) kromosoomistikus puudub mingi kromosoom täielikult. Tõenäoliselt kõigil liikidel on nullisoomne kromosoomistik varases embrüogeneesis letaalne, kuid sugurakkudes (spoorides) võib see esineda.
ontogenees (ontogeny) -- organismi (isendi) individuaalne areng alates viljastumisest ja lõpetades surmaga. Jaguneb embrüogeneesiks e. prenataalseks arenguks ja postembrüogeneesiks e. postnataalseks arenguks; viimases eristatakse tavaliselt kolm järku: nooriga, täisiga ja raugaiga; osal loomadel kulgeb nooriga moondega. Varastel etappidel kordab o. mõningaid fülogeneetiliste eellaste arengufaase.
panmiksis (panmixis) -- e. vaba ristumine (random mating), populatsioonile omane ristumissüsteem, kus iga indiviid võib paaruda mis tahes genotüüpi omava vastassoost indiviidiga ning erinevate genotüüpidega indiviidide ristumiste sagedus oleneb ainult nende sagedusest populatsioonis (st. on juhuslik sündmus). Panmiksist hinnatakse tavaliselt üksikute lookuste suhtes eraldi. Kui populatsioonis on mingi autosoomse lookuse järgi kolm genotüüpi AA, Aa, aa sagedustega vastavalt D, H ja R, siis on panmiksise puhul 6 erinevat ristumistüüpi sagedustega (D + H + R)2 = D2 + H2 + R2 +2DH + 2DR + 2HR.
penetrantsus (penetrance) -- sagedus (%), millega mingi konkreetne genotüüp avaldub selle kandjate fenotüübis. Tavaliselt kasutatakse seda mõistet mingi dominantse mutantse alleeli avaldumissageduse hinnanguna heterosügootide hulgas. Täieliku penetrantsuse korral avaldub kõigil heterosügootidel vastav mutantne tunnus (puue, haigus), mittetäieliku (e. osalise) penetrantsuse puhul aga ainult teatud osal (nt. 90%) vastava genotüübiga indiviididest. Penetrantsus sõltub nii genotüübilisest taustast (nn. modifikaatorgeenidest) kui ka keskkonnatingimustest, milles indiviidid arenevad ja elavad.
perekondlik (familial) -- perekondades toimuv, perekonniti esinev, teatud perekondadele iseloomulik tunnusevariant või haigus.
pleiotroopsus (pleiotropy) -- vt. polüfeensus.
ploidsus (ploidy) -- samatüübiliste (homoloogiliste) kromosoomide (või liigiomaste kromosoomikomplektide) kordsus indiviidi (raku) kromosoomistikus. Enamiku eukarüootsete organismide elutsüklis vahelduvad kahekordselt erinevad ploidsusfaasid -- gameetne haplofaas ja sügootne diplofaas. Vt. monoploidsus, haploidsus, diploidsus, polüploidsus, euploidsus, aneuploidsus, autoploidsus,
alloploidsus.
polüalleelsus (polyallelism) -- e. mitmealleelsus (multiple allelism), nähtus, mil uuritaval geenil (või muul geneetilisel markeril) on samas populatsioonis enam kui kaks alleeli. Vt. nt. AB0 vererühmasüsteem. Polüalleelsuse korral on panmiktilises populatsioonis vastava lookuse puhul erinevaid genotüüpe [k*(k+1)]/2, kus k on geeni alleeliarv.
polüfeensus (polypheny) -- e. pleiotroopsus (pleiotropy), ühe (mutantse) geeni samaaegne fenogeneetiline toime mitmele erinevale tunnusele. Harvadel juhtudel tuleneb polüfeensus sellest, et ühe geeni produktideks võib olla kaks või enam iseseisva funktsiooniga polüpeptiidi (või peptiidhormooni). Enamasti on polüfeensus tingitud ühe geeni produkti osalemisest erinevates ainevahetusreaktsioonides või mitmetes rakulistes vastasmõjudes ja arenguprotsessides. Polüfeense toimega geeni mutatsioon avaldub tavaliselt mitme tunnuse üheaegse muutusena, nt. patoloogia korral sündroomina.
polügeenid (polygenes) -- mitme lookuse geenid, mille alleelsed kombinatsioonid mõjutavad sama tunnuse avaldumisastet. Alleelsed asendused üksikutes lookustes põhjustavad väikesi erinevusi indiviidide fenotüüpides; selliste geenide avaldumine sõltub olulisel määral ka keskkonnatingimustest. Seetõttu on neid geene ja individuaalseid genotüü on raske tuvastada. Enamasti on sellised geenid polümeerses (aditiivses) vastasmõjus.
polügeensus (polygeny) -- 1) üldisemas tähenduses: tunnuse determineeritus mitme geeni koostoimest, kusjuures geenitoimete vahel võivad olla erinevad interaktsioonid. 2) kitsamas tähenduses: polümeersus (eriti inglise terminoloogias).
polümeersus (polymery) -- vt. polümeerne geenitoime.
polümeerne geenitoime (polymeric gene action) -- e. aditiivne geenitoime, kahe või (enamasti) mitme geeni samasuunaline ja kumuleeruv toime ühele tunnusele. Selline geenide koostoime põhjustab tunnuse erinevaid avaldumistasemeid, kvantitatiivse varieeruvuse erinevate mõõdetavate tunnuseväärtuste vahel. Igal polümeersüsteemi geenil eristatakse tunnust intensiivistavaid pluss-alleele ja mittetoimivaid null-alleele. Tunnuseväärtus oleneb eri lookuste pluss-alleelide koguarvust (geenidoosist), kuid mitte sellest, milliste geenide pluss-alleelid genotüübis on (genotüübid AAbb, aaBB ja AaBb on fenotüübiväärtuse poolest samaväärsed). Kui tunnust kujundavad 4-5 või enam polümeerset geenipaari, siis ilmneb populatsioonis tunnuse pidev (enamasti normaaljaotusele alluv) muutlikkus. Seda soodustab asjaolu, et polümeersed tunnused alluvad olulisel määral samasuunalistele keskkondlikele mõjutustele (modifikatsioonidele). Polümeersete tunnuste geneetilise analüüsiga tegeleb nn. biomeetriline geneetika.
Ingliskeelses traditsioonis samastatakse polümeersus tihti polügeensusega (vt. polügeenid ja polügeensus).
polüploidsus (polyploidy) - indiviidi (raku) kromosoomikomplektide paljukordsus; haplofaasis (gameetides) on kromosoomikomplekte rohkem kui üks (n > x) ja diplofaasis (sügootides) rohkem kui kaks (2n > 2x) (Vt. ploidsus). Polüploidsuse astet määratletakse diplofaasi (so. sügootse) kromosoomistiku kordsuse kaudu: tri-, tetra-, heksa-, oktaploidsus jne. Polüploidsus on laialt levinud taimeriigis, loomariigis harva. Inimesele on polüploidsus reeglina letaalne, leitud peamiselt aborteerunud loodetest, elusaltsündinud lastel üliharva triploidsust.
Nähtust, kus ainult osa indiviidi (mõne koe või organi) rakkudest on polüploidsed, nim. polüsomaatiaks. See on paljudel organismidel (ka inimesel) normaalne nähtus.
polüsomaatia (polysomatia) -- nähtus, kus kogu organismi või tema mingi koe koosseisus on kõrvuti diploidsete rakkudega ka polüploidseid. Polüsomaatia on küllalat levinud nähtus, tavaliselt omane kõrge funktsionaalse koormusega organite kudedele (inimesel nt. peamiselt maksas ja südamelihases). Polüploidsed rakud võivad tekkida rakkude ja tuumade ühinemise teel, kuid enamasti endomitoosi ja endoreduplikatsiooni kaudu.
polüteenne kromosoom (polytene chromosome) -- polüteenkromosoom e. hiidkromosoom (giant chromosome), paljuniidiline kromosoom, interfaasikromosoomide eritüüp, mis on tekkinud korduvalt endoreduplitseerunudkromatiidide lahknematuse tulemusena. Reduplikatsioonide arv polüteensete kromosoomide arengus võib ulatuda 10...12, seega kromatiidide arv ühes kromosoomis võib olla vahemikus 1024...4096. Üksikkromatiidid on tihedalt koos, nii et selline kromosoom näib tervikmoodustisena ja on valgusmikroskoobi abil hästi vaadeldav interfaasi tuumas; nad on 200 või isegi enam kordi pikemad kui meioosis ja mitoosis vaadeldavad kromosoomid. Osal juhtudel (Diptera süljenäärmetes) on homoloogilised polüteenkromosoomid liibunud teineteisega kokku (konjugeerunud), mis veelgi suurendab nende läbimõõtu. Polüteenkromosoomidele on iseloomulik vöötjas värvumine -- piki kromosoomi vahelduvad kitsad, kuid siiski varieeruva laiuse ja tihedusega tumedad vöödid (bändid e. diskid) heledamate vöötidega. Tumedad vöödid vastavad tihedalt liibunud kromatiidide identsetele kromomeeridele. Aktiivsete geenide või geeniklastrite piirkonnad avalduvad polüteenses kromosoomis väiksemate või suuremate rõngaspaksendite e. puhvidena (puffs), mille eriti suuri vorme nim. Balbiani rõngasteks (Balbiani rings). Neis toimub intensiivne RNA süntees. Indiviidi arengu eri etappide ja eri kudede rakkudes puhvide ja Balbiani rõngaste lokalisatsioon muutub; see nähtus oli esimeseks tõendiks geenide valikulise aktivatsiooni kohta ontogeneesis ja rakkude diferentseerumisel. Klassikaliselt tuntud polüteenkromosoomid on kahetiivaliste putukate (Diptera) vastsete süljenäärmerakkudes, kuid esinevad ka nende ja mõnede teiste putukate vastsete ja valmikute muudes kudedes, osal protistidel ja õistaimedel. Polüteenkromosoomide uurimine on olnud suure tähtsusega kromosoomide kaardistamisel ja arengugeneetiliste probleemide selgitamisel.
polüteensus (polyteny) -- osa organismide teatud rakutüüpide eriline tuumadiferentseerumise nähtus, mida iseloomustab polüteenkromosoomide olemasolu.
populatsioonigeneetika põhiseadus (population genetics' basic law) -- vt. Hardy-Weinbergi seadus.
populatsiooni geneetilise tasakaalu seadus (the law of population's genetic equilibrium) -- vt. Hardy-Weinbergi seadus.
prokarüoot (prokaryote) -- eeltuumse e. prokarüootse rakuehitusega organism. Sellise rakuehitusega organismid on (eu- ja arhe-)bakterid (sh. nn. sinivetikad e. sinikud, Cyanobacteria).
prokarüootne rakk (prokaryotic cell) -- e. eeltuumne rakk, evolutsiooniliselt algsem ja primitiivsem rakuehituse tüüp kui eukarüootne e. päristuumne rakk. Prokarüootne rakutüüp on omane bakteritele. Prokarüootne rakk on võrreldes eukarüootsega palju väiksem (läbimõõduga enamasti vahemikus 0,1...5 mikromeetrit); tema geneetilised elemendid paiknevad tsütoplasmas (pole membraanidega piiristatud tuuma); genoomiks on üks rõngas-DNA molekul (osal juhtudel väikesed rõngasjad lisa-DNA molekulid -- plasmiidid), mis pole püsivalt seotud valkudega (histoonid puuduvad); rakus puuduvad membraansed organellid (mitokondrid, plastiidid, tsütoplasmavõrgustik, Golgi kompleks jms.).
pseudoautosoomsus (pseudoautosomy) - geenide paiknemine X- ja Y-kromosoomi homoloogilises regioonis (PAR), mille vahel võib toimuda ristsiire. See regioon moodustab väikese osa (inimesel ca 2,6 Mb) nende kromosoomide lühikese õla tipust. Need geenid on mõlemal sugupoolel paarilised ja päranduvad samal viisil kui autosoomsed geenid (kuid asuvad gonosoomides). Neid geene ja neist määratud tunnuseid nimetatakse samuti pseudoautosoomseteks (inimesel nt. MIC2 ja XGR).
pudelikaela efekt (bottleneck effect) -- ajutise e. mööduva geneetilise triivi üks erijuhte, mis tuleneb populatsiooni arvukuse järsust, kuid 1...3 põlvkonna jooksul mööduvast arvukuse langusest. Populatsiooni arvukuse suur langus ja järgnev lühiajaline madalarvukuse ("pudelikaela") periood põhjustavad olulisi juhuslikke muutusi geenifondis. Need muutused on oma olemuselt sarnased asutaja efektiga, selle erinevusega, et ei teki emapopulatsioonist eraldatud uut populatsiooni, vaid algselt suur populatsioon ise teeb läbi ajutise arvukuse madalseisu ja taastub seejärel samal asualal, kuid suuresti juhuslikult muutunud geneetilise struktuuriga. Populatsiooni arvukuse kollaps võib olla tingitud looduslikest katastroofidest, kuid osal loomadel esinevad perioodiliselt korduvad arvukuse järsud langused ja tõusud -- nn. populatsioonilained e. elulained.
punktmutatsioon (point mutation) -- lokaalne geenmutatsioon, geenisisene mutatsioon, mis puudutab üksikuid koodoneid ja võib rekombineeruda sama geeni teiste punktmutatsioonidega (erinevaid mutantseid alleele sisaldavates heterosügootides).
pärandumine (inheritance) -- geneetiliste struktuuride (kromosoomide, geenide) ja neis sisalduva geneetilise informatsiooni ülekanne vanematelt järglastele (või emarakult tütarrakkudele), mille tulemusena kujunevad viimaste genotüübid. Pärandumise seaduspärasused sõltuvad organismide sigimis- ja rakkude jagunemisviisist. Mittesugulisel sigimisel (ja rakkude mitootilisel jagunemisel) pärandub vanema genotüüp tervikuna kõigile järglastele, mille tulemusena tekib geneetiliselt identne järglaskond (kloon). Sugulisel sigimisel (meioosi ja viljastumise vahendusel) toimub erinevatelt vanematelt pärit geneetiliste elementide lahknemine ja kombineerumine, nii et järglased on olulisel määral erinevate genotüüpidega. Selline pärandumine allub Mendeli ja Morgani seadustele. Kromosomaalse soomääramisega organismidel sõltub geneetiliste elementide (geenide) pärandumise viis veel nende lokalisatsioonist kas autosoomides või sugukromosoomides.
Iseäraliku pärandumisviisiga on tsütoplasma geneetilised elemendid (mitokondrite ja kloroplastide genoomid). Need päranduvad üksnes munaraku kaudu (emapoolne pärandumine), neil ei toimu rekombinatsiooni, ning geneetiline lahknemine on reeglipäratu (kusjuures harvaesineva generatiivse lahknemise kõrval ilmneb sagedam somaatiline lahknemine).
Sageli kasutatakse mõistet pärandumine ka tunnuste kohta, kuigi tunnused otseselt ei pärandu, vaid kujunevad genotüübi avaldumise tulemusena fenogeneesis.
pärilikkus (heredity) -- üks elu põhiomadusi, bioloogilise spetsiifika järjepidevus organismide, rakkude ja viiruste paljunemisel ning organismide ja rakkude eneseuuendamisel. Pärilikkus on informatsiooniline nähtus, tema sisuks on biostruktuure ja -funktsioone määrava instruktiivse informatsiooni säilitamine, edastamine ja avaldumine. Pärilikkus on kompleksnähtus; seda võib käsitleda ja uurida substraadi e. replitseeritavate geneetiliste struktuuride tasemel (genoomid, kromosoomid, DNA ja RNA, geenid) ning funksionaalsete protsesside aspektis (pärandumine: genotüüp --> genotüüp; fenogenees: genotüüp --> fenotüüp). Pärilikkus sisaldab endas nii pärilikku konstantsust (püsivust) kui ka pärilikku muutlikkust (mutageneesi, rekombinatsiooni) tagavad mehhanismid. Pärilikkust ja sellega seotud nähtusi uurib geneetika.
päritavus (heritability) -- e. heritaablus, biomeetrilise geneetika mõiste: kvantitatiivse tunnuse populatsioonisisese muutlikkuse see osa, mis on tingitud pärilikest (st. genotüübilistest) erinevustest indiviidide vahel; ülejäänud osa tunnuse varieeruvusest on tingitud kas puhtalt eksogeensetest (keskkonna-) tingimustest või genotüüpide ja keskkonnategurite vastasmõjust ning kovariatsioonist. See on päritavuse määratlus laiemas mõttes (broad-sense heritability). Päritavus kitsamas mõttes (narrow-sense heritability) tähendab üksnes aditiivsest e. polümeersest geenitoimest tingitud fenotüübiliste erinevuste osakaalu. Päritavuse arvuliseks hinnanguks on päritavuskoeffitsient, mida määratakse biomeetrilise geneetika meetoditega.
päritavuskoefitsient (heritability coefficient) -- tähis: h2; arvuliselt väljendatav geneetilise muutlikkuse suhtosa tunnuse üldises populatsioonisiseses (individuaalses) muutlikkuses antud keskkondlikes tingimustes. Üldjuhul defineeritakse geneetilise dispersiooni suhtena tunnuse kogudispersiooni: h2 = Vg/Vp. Päritavuse sisu õigeks mõistmiseks on vaja arvestada, et päritavuskoefitsient hindab tunnuse geneetilise muutlikkuse osa antud geneetilise struktuuriga populatsioonis konkreetsetes keskkonnatingimustes; see ei näita tunnuse päriliku tingituse määra ega mehhanismi üksikindiviidide arengus.
rajaja efekt (founder effect) -- ka asutaja efekt; geenitriivi üks mehhanisme, mis seisneb selles, et emapopulatsioonist irdunud üksikud isendid (või indiviidipaarid) panevad aluse uuele populatsioonile, mille geneetiline struktuur erineb oluliselt lähtepopulatsiooni omast. Seda mõistet kasutatakse ka juhul, kui mingis populatsioonis levib teatav geneetiline spetsiifika (nt. mingi mutatsioon), mis on saanud alguse ühest (või mõnest) immigrandist, kes on mingil põhjusel olnud keskmisest kõrgema sigivusega.
reaktsiooninorm (reaction norm) -- pärilik reaktsiooninorm, modifikatsiooniliste muutuste sõltuvus isendi genotüübist; tunnusevariantide ja nende avaldumisastmete kogum, mis võib sama genotüübiga organismide (rakkude) fenotüübis ilmneda nende arengul ja elutegevuses erinevates keskkonnatingimustes. Reaktsiooninorm määrab antud genotüübiga indiviidi modifikatsioonide kvalitatiivsed ja kvantitatiivsed piirid, st. selle, millist laadi ja missuguse ulatusega (amplituudiga) tunnusemuutused võivad mingite keskkonnamuutuste (või isendi aktiivsuse muutuste) korral aset leida.
rekombinant (recombinant) -- iga indiviid või rakk, mis on tekkinud vanemtüüpi genotüüpide (fenotüüpide) ümberkombineerumise (geneetilise rekombinatsiooni) tagajärjel. Vastandatakse vanemtüübipidele. Rekombinantide tekkesageduse määramine analüüsival di- või trihübriidsel ristamisel on geeniahelduse ja ristsiirde uurimise aluseks.
retsessiivsus (recessivity) - ühe tunnuse (alleeli) varjuvus tunnusepaaris (alleelipaaris) heterosügootse genotüübi puhul; tunnuse (sh. defekti, haiguse) avaldumine ainult homosügootsuse (aa) korral. Retsessiivsus on dominantsuse vastandnähtus.
Rhesus-süsteem (Rhesus system) -- Rh-süsteem, inimese vererühmasüsteem, milles erinevused võivad põhjustada komplikatsioone vereülekandel ning ema ja loote immunoloogilisi konflikte. Meditsiinilises käsitluses ("Rh-faktor") eristatakse tavaliselt Rh-positiivset ja Rh-negatiivset rühma (eurooplastest on ca 16% Rh-negatiivsed); esimesel juhul on erütrotsüütide pinnal immunoloogiliselt aktiivne valgulise olemusega Rh-D antigeen, teisel juhul see puudub. Tegelikult on Rh-süsteem palju heterogeensem, on eristatud üle 20 Rh-fenotüübi. Rh-süsteemi geneetiline olemus oli kaua aega ebaselge; alates 1944. a. konkureerisid 2 geneetilist mudelit. A. Wiener'i hüpoteesi kohaselt oli süsteemi heterogeensus määratud ühe polüalleelse geeni poolt (allelid R0, R1, R1W, R2, Rz, r, r', r" ja rida haruldasemaid). A. Race'i ja R. Fisher'i hüpoteesi järgi olid Rh-fenotüübid määratud 3 tihedalt aheldunud geeni (C,D,E) poolt, millest igaühel eristati 2 või 3 alleeli (tavalisemad haplotüübid: CDe, cDE, cDe, CWDe, CDE, cde, Cde, cdE). Alles 1991. a. sai süsteemi geneetiline loomus tegeliku molekulaargeneetilise seletuse. Kromosoomiregioonis 1p36 asub 2 kõrvuti paiknevat homoloogilist geeni: RHD- ja RHCE. D-geeni dominantne alleel kodeerib D-valku (D-antigeeni), retsessiivne alleel on defektne (ei anna funktsionaalset valku). CE-geen kodeerib alternatiivse splaissingu vahendusel 2 erinevat alleelse polümorfismiga valku (Cc- ja Ee-variandid).
ristsiire (crossing over) -- homoloogsete aheldusrühmade (kromosoomide) paarides toimuv retsiprookne segmendivahetus, mis põhjustab intrakromosoomse geneetilise rekombinatsiooni (alleelide vahetuse) aheldunud lookuste vahel. Eukarüootsetel organismidel toimub ristsiire meioosi profaasis konjugeerunud homoloogsete kromosoomide kromatiidiosade vahetusena. Ristsiirde sagedus kahe lookuse vahel sõltub nendevahelisest kaugusest piki kromosoomi (vt. Morgani seadus).
sagedusest sõltuv valik (frequency-dependent selection) -- loodusliku valiku vorm, mille puhul fenotüübi (~genotüübi) kohasus sõltub tema sagedusest populatsioonis: kohasus on pöördvõrdelises seoses fenotüübi sagedusega. Kui fenotüüp on haruldane, siis on ta eeliskohasusega; kui selle sagedus ületab teatud optimaalse taseme, siis on ta valikuliselt ebasoodus (võrreldes alternatiivsete tunnustega). Selline valik loob ja säilitab polüalleelset adaptiivset polümorfismi populatsioonides.
sugukromatiin (sex chromatin) -- e. Barri kehake, emasimetajate (sh. naiste) keharakkude tuumas paiknev kromatiinivärvidega tugevalt värvuv kehake (läbimõõduga ca 1 µm), mis tavaliselt asub tuuma perifeerias tihedalt vastu tuumamembraani. Sugukromatiini olemasolu/puudumise järgi saab väheste interfaasirakkude (nt. suuõõne limaskestakaape või juuksekarva juurenääpsu preparaadi) mikroskoopilisel uurimisel tuvastada indiviidi kromosoomset (st. geneetilist) sugu (vt. gonosoom). X-kromosoomide arvu anomaaliate avastamise järel leiti, et sugukromatiinkehakeste arv on alati ühevõrra väiksem X-kromosoomide arvust karüotüübis: Turneri sündroomiga naistel (karüotüüp 45, X) puudub nagu normaalsetel meestelgi (46, XY), Klinefelteri sündroomiga meestel (47, XXY) üks nagu normaalsetel naistel, X-trisoomiaga naistel (47, XXX) kaks jne. Sugukromatiinkehakesed kujutavad endast kondenseerunud (inaktiveeritud) X-kromosoome -- imetajate somaatilistes rakkudes on aktiivseisundis ainult üks X-kromosoom. Vt. ka geenidoosi kompensatsioon.
sugukromosoom (sex chromosome) -- vt. gonosoom.
suguliitelisus (sex-linkage) - geenide ja nendest määratud tunnuste sooseoselisus, mis on tingitud nende geenide lokalisatsioonist sugukromosoomide X- ja Y-spetsiifilistes (mittehomoloogsetes) piirkondades. Suguliitelisi geene pärandavad sugupooled oma eri sugu järglastele erinevalt (retsiprooksed ristamised annavad erineva järglaskonna). Suguliitelised tunnused esinevad sugupooliti erineva sagedusega või on piiratud ühe sugupoolega. Enamasti mõistetakse suguliitelisuse all ainult X-liitelisust, seejuures Y-liitelisust tähistatakse terminiga holandrilisus. Suguliitelisuse vastandnähtus on autosoomsus.
sugupuu (pedigree) - suguvõsa (perekonna) genealoogiline skeem, millele on tingmärkide abil kantud iga indiviidi sugu, eellased, järglased ja külgsugulased võimalikult paljude põlvkondade ulatuses; geneetilise uurimise korral märgitakse ka vaatlusaluse tunnuse (haiguse) esinemine. Sugupuid kasutatakse geneetilisel uurimisel genealoogilise meetodi abil, eriti inimesegeneetikas.
sõltumatusseadus (law of independent assortment, law of independence) -- e. sõltumatu lahknemise seadus e. vaba kombineerumise seadus, Mendeli III seadus.
triploidsus (triploidy) -- indiviidi (raku) kromosoomistiku kolmekordsus (st. indiviidi kromosoomistikus esineb 3 kromosoomikomplekti, va. gonosoomid, mille suhtes võivad esineda erinevad arvsuhted).
ulukalleel (wild allele) -- e. metsalleel, looduslikes populatsioonides mingi geeni peamine alleel (nö. standardalleel), vastandatuna harvadele mutantsetele alleelidele. Polümorfses populatsioonis võib olla mitu ulukalleeli. (See eestikeelne termin on seni kasutusel olnud loomageneetikas).
vaba ristumine (random mating, free mating) -- vt. panmiksis.
valik (selection) -- genotüüpide ebavõrdne ja mittejuhuslik reproduktsioon sõltuvalt nende erinevast kohasusest mingi välise (valiku-) teguri suhtes. Bioloogiline valik võib toimuda elu organisatsiooni erinevatel tasemetel (geneetiliste rakuorganellide, gameetide ja hulkrakse organismi rakkude, indiviidide ning indiviidirühmade (deemide) vahel). Valiku tagajärjel muutuvad geenide (alleelide) ja genotüüpide sagedused valikuobjektide populatsioonides. Valik oleneb genotüüpide erinevast toimest kas eluvõimele või paljunemisele. Valikul on nii positiivne kui negatiivne aspekt: ühtede genotüüpide eelistatud üleelamine (survival) ja reproduktsioon käib koos teiste genotüüpide (suhtelise) eliminatsiooniga. Eristatakse looduslikku ja kunstlikku valikut.
vanemtüüp (parental type) -- hübridoloogilisel analüüsil hübriidide järglaskonnas lahknevate vormide hulgas üks kahest genotüübist (fenotüüist), mis vastab ühele või teisele ristatud pärivuspuhastest vanemvormidest (tõugudest, sortidest, liinidest). Vastandatakse rekombinantidele.
vererühmasüsteem (blood group system) -- e. veregrupisüsteem, teatud tüüpi antigeenide variandid erütrotsüütide pinnal, mille suhtes populatsiooni indiviidid jagunevad erinevatesse rühmadesse. Need variandid (rühmad) on määratud ühe geeni alleelide, mõnikord aga tihedalt aheldunud lookuste haplotüüpidepoolt. Inimese tuntumad polümorfsed vererühmasüsteemid on AB0, Rhesus, MNSs, P, Kell, Lewis, Lutheran, Duffy, Kidd, Diego ja Xg.
W-kromosoom (W-chromosome) -- emaassoo heterogameetsuse korral sageli kasutatav tähis sellele gonosoomile, mis esineb ainult emassool (isassoo heterogameetsuse puhul vastab sellele Y-kromosoom).
õved (siblings, sibs) -- ühe vanemapaari järglased; õed-vennad koosvaadatuna (ains. õvi). Võib eristada täisõvesid (fullsibs) ja poolõvesid (halfsibs).
ühetaolisuse seadus (law of uniformity) -- e. ühtlikkuse seadus, Mendeli I seadus.
üledominantsus (overdominance) -- üledomineerimine, populatsioonigeneetiline nähtus, mille puhul heterosügoodid mingi geeni suhtes on suurema kohasusega (valikueduga) kui vastavad homosügoodid (AA<Aa>aa). Sel juhul eelistab valik heterosügoote (ja elimineerib homosügoote), mis viib tasakaalulise adaptiivse polümorfismi tekkele ja säilimisele populatsioonis. Klassikaline näide on polümorfism sirprakulisuse hemoglobiini suhtes troopilise malaaria alade inimpopulatsioonides (heterosügoodid on suurema resistentsusega malaariale kui normaalsed homosügoodid).
Xg -süsteem (Xg system) -- ainus X-liitelinevererühmasüsteem inimesel. Süsteem on dialleelne, alleelid Xga ja Xg, fenotüübid (vererühmad) Xg(a+) ja Xg(a-); Xg(a+) on naistel dominantne. Euroopa elanikkonnas on u. 88% naistest ja 65% meestest Xg(a+). Xg-lookus paikneb Xp2-regioonis, see ei allu laionisatsioonile; tema avaldumistaset reguleerib pseudoautosoomne geen XGR.
X-kromosoom (X-chromosome) - üks gonosoomidest, esineb homogameetse sugupoole (nt. normaalse emasimetaja, ka naise) kromosoomistikus paariliselt, heterogameetse sugupoole (nt. isasimetaja, ka mehe) kromosoomistikus paaritult. Inimese X-k. on suuremapoolne submetatsentriline kromosoom; selles on lokaliseeritud üle 300 geeni. Vrd. Y-kromosoom.
X-liitelisus (X-linkage) - (suguliitelisus kitsamas mõttes), mingi geeni ja sellest määratud tunnuse (sh. defekti, haiguse) sooseoselisus, mis on tingitud selle geeni paiknemisest ainult X-kromosoomis. X-liitelised geenid on emassool paarilised, isassool paaritud (hemisügootsus). Nende pärandumisiseärasused seisnevad selles, et ema pärandab neid kõigile järglastele ühtmoodi, isa aga ainult tütardele. Retsessiivsed X-liitelised defektid esinevad peamiselt isasool (meestel), kuigi neid põhjustavad alleelid päranduvad enamasti heterosügootsete emaste (naiste; nn. konduktorite) kaudu. Inimesel on X-liitelised tunnused nt. Xg-vererühmad, puna-rohe-värvipimedus (daltonism), Duchenne'i lihasdüstroofia, hemofiilia A ja B jm. Vrd. autosoomsus, Y-liitelisus.
Y-kromosoom (Y-chromosome) - üks gonosoomidest, esineb ainult heterogameetsel sugupoolel (nt. isasimetajatel, ka mehel) koos ühe X-kromosoomiga. Sisaldab tavaliselt peale heterogameetsele sugupoolele spetsiifiliste (Y-liiteliste) geenide ka X-kromosoomiga homoloogse (pseudoautosoomse) regiooni. Imetajate Y-kromosoomis lokaliseeruv geen SRY määrab valgulise testiseid determineeriva faktori (TDF) kaudu embrüo (gonaadide) isasarengu. Inimese Y-kromosoom on väike akrotsentriline (kuid satelliidita) kromosoom; selles on identifitseeritud ca 30 geeni.
Y-liitelisus (Y-linkage) - suguliitelisuse vorme, mingi geeni või muu geneetilise markeri lokaliseeritus Y-kromosoomis (X-kromosoomiga mittehomoloogilises osas) ja selle pärandumine ainult Y-kromosoomi kaudu (heterogameetse isassoo puhul piki isasliini, holandriliselt). Imetajatel on Y-liitelised testiseid determineeriva faktori (TDF) geen (SRY), spermatogeneesi kontrollivad ja testisespetsiifilisi valke määravad geenid, isaspetsiifilise H-Y antigeeni sünteesi reguleeriv geen, aga ka vähemalt üks kasvu kontrolliv geen (GCY). Vrd. X-liitelisus.
Z-kromosoom (Z-chromosome) -- emassoo heterogameetsuse korral sageli kasutatav tähis sellisele gonosoomile, mis esineb mõlemal sugupoolel (isasheterogameetsuse puhul vastab sellele X-kromosoom).