Geneetiline mitmekesisus
Geneetiline mitmekesisus tähendab pärilikke molekulaarseid erinevusi populatsiooni sees või populatsioonide vahel.
Enamasti mõistetakse geneetilise mitmekesisuse all liigisiseseid DNA järjestuse erinevusi, aga varieeruvust saab võrrelda ka liikide vahel. Geneetiline mitmekesisus kui evolutsiooniliste muutuste alus on alati olnud põhiküsimus evolutsioonilises geneetikas, aga ka põllukultuuride ja koduloomade aretamises ning meditsiinis. Samuti on geneetiline mitmekesisus kesksel kohal looduskaitsegeneetikas, mille eesmärk on tagada liikide kohastumisvõime muutuvas keskkonnas. Hoolimata olulisest rollist evolutsiooniteoorias ja avastustest molekulaarbioloogias, on geneetilises mitmekesisuses ja selle püsimises veel palju mõistatuslikku.
Uurimise ajalugu
Molekulaarse evolutsiooni ajastu juhatasid sisse kolm peamist avastust:
- Clement Markert ja Freddy Moller leidsid 1959. aastal ensüümide molekulaarse mitmekesisuse ja nende tähtsuse geneetikas, füsioloogias, arengus ja evolutsioonis;
- Emile Zuckerkandl ja Linus Pauling identifitseerisid 1965. aastal valkude järjestuste varieeruvuse liikide vahel, mille tulemusena postuleeriti molekulaarne kell;
- 1966. aastal hindasid Richard Lewontin, John Lee Hubby ja teised ensüümide varieeruvuse äädikakärbsel ja Harry Harris inimesel.
Molekulaarbioloogilised saavutused võimaldasid iseloomustada indiviidide, populatsioonide ja liikide geneetilist mitmekesisust. Esmalt selgusid suhted geenide ja valkude vahel, edaspidi tuumasisesed ja tuumavälised kodeerivad ja mittekodeerivad DNA regioonid, geenide, geenidevaheliste alade (intergenic spacers) ja geeniperekondade struktuur, avaldumine, funktsioon, mehhanismid ning evolutsioon. Selleks kasutati rekombinantse DNA meetodeid.
Edasijõudmised molekulaarsetes uuringutes leidsid aset tänu uute laboratoorsete meetodite kasutuselevõtule. Mõjukaimad tehnoloogiad on olnud: valkude elektroforees 1960ndate lõpus ja 1970ndatel,rakutuuma ja mitokondriaalse DNA restriktsioonifragmentide pikkuse polümorfismi (RFLP) analüüsid 1970ndate lõpul ja 1980ndatel, DNA sõrmejäljed 1980ndate keskpaigast lõpuni ja DNA sekveneerimine polümeraasi ahelreaktsiooni (PCR) abil 1990ndatel.
Võrdleva genoomika ja genoomiuuringute üks eesmärke on juhtida tähelepanu evolutsiooniliste muudatuste ning bioloogilise info avaldumise, tähenduse, edastamise ja regulatsiooni geneetilistele alustele.
Geneetilise mitmekesisuse tähtsus
Geneetilise mitmekesisuse funktsiooni populatsioonis on selgitanud paljud uuringud. Rikkalik genofond pakub looduslikule valikule materjali, et valida paremini kohastunud genotüüpe. Keskkonna muutumise korral võimaldavad variatsioonid geenides genereerida muutusi organismi fenotüübis, et muuta organismi kohasemaks uutes oludes. Populatsioonides, kus on kõrge geneetiline mitmekesisus, on rohkem alleelivariante, mille hulgast saab looduslik valik sagedasemaks muuta kõige sobivamad. Seega on geneetiline mitmekesisus tähtis evolutsiooni toimumiseks. Madala geneetilise mitmekesisusega liikidel on suurem risk välja surra. Madal geneetiline mitmekesisus ei pruugi põhjustada ainult populatsiooni kohastumisvõime langust, vaid ka tõsta energeetilisi kulutusi ning tuua kaasa homöostaasi ja produktiivsuse vähenemise. Geneetiline mitmekesisus tagab geneetiliste distantside esinemise liigisiseselt ja liikide vahel. Teadmised sellest teevad võimalikuks liikide ja tüvede identifitseerimise geneetilise profiili alusel, fülogeneesipuude rajamise, on abiks looduskaitse majandamisel, põllumajanduses ja meditsiinis.

Näiteid geneetilise mitmekesisuse ökoloogilisest olulisusest
- Meemesilaste kolooniates avaldab geneetiline mitmekesisus positiivset mõju. Kõrge mitmekesisusega kolooniad säilitavad vastusena temperatuuri muutustele edukamalt ühtlast temperatuuri kui need, kus mitmekesisus on madal. Ilmselt on põhjuseks tööliste erinevused geenides, mis määravad temperatuuri, millest alates hakkab neil avalduma vastav käitumine pesatuulutamiseks.
- Põlluviljade monokultuurides on taimed geneetiliselt väga sarnased ja see suurendab nende vastuvõtlikkust haigustekitajatele. Kui peremeest patogeeni vastu kaitseva alleeli sagedus on kõrge, siis juhul, kui patogeen muutub võimeliseks kaitsest läbi murdma, saab ta edukalt levida. Näiteks 1840ndatel Iirimaad laastanud näljahäda oli põhjustatud kartulite epideemiast. Kartulisorti „lumper“ nakatas mädanikku põhjustav protist Phytophthora infestans.
- Geparditel on võrreldes teiste suurte kaslastega madalam reproduktiivsus ja suurem vastuvõtlikkus haigustekitajatele. Selle põhjuseks arvatakse olevat madal geneetiline mitmekesisus. Molekulaarsete analüüside tulemused näitasid, et gepardid on läbinud kaks populatsiooni pudelikaela. Esimene kord 10 000 aastat tagasi ja teine viimase 100 aasta jooksul.
Geneetilise mitmekesisuse võimalikud allikad
Protsessid, mis põhjustavad geneetilist varieeruvust:
- Geneetilised mutatsioonid
- Homoloogiline rekombinatsioon suguliselt paljunevate organismide meioosis, kui kaks homoloogilist kromosoomi vahetavad võrdse pikkusega osi ning tekivad rekombinantsed kromosoomid.
- Horisontaalne geeniülekanne, mis esineb sagedasti näiteks bakteritel. Bakterid saavad DNA-d omastada väliskeskkonnast transformatsiooni teel, teistelt bakteritelt kas viiruste vahendatud transduktsiooni või bakterirakkude vahelist kontakti eeldava konjugatsiooni teel.
- Polüploidsus – organismil on rakus homoloogilisi kromosoome rohkem kui kaks. Seega on ka ristsiirde tulemusena tekkiv geneetiline varieeruvus suurem.
Geneetilise mitmekesisuse säilimine looduses
Geneetilise mitmekesisuse säilimine looduslikes populatsioonides on olnud evolutsioonilise geneetika keskne probleem alates valkude polümorfismi avastamisest. Nüüdseks on teada, et ainult juhuslikkusel põhinevad seletused, mis seovad heterosügootsust peamiselt populatsiooni suuruse efektidega, ei ole realistlikud. Populatsioonid on abiootiliste (nt kliima) ja biootiliste (nt. parasiidid, patogeenid, konkurents) faktorite tõttu harva tasakaaluseisundis. Seetõttu esineb ka dünaamiliselt muutuvate looduslike populatsioonide geneetilises mitmekesisuses järske tõuse ja langusi ökoloogilise surve mõjul. Peamised mudelid selgitavad geneetilise mitmekesisuse püsimist looduses pigem seoses keskkonna heterogeensuse (erinevad nišid) ja survega kui efektiivse populatsiooni suuruse ja geenivooluga. Looduslik valik tundub olevat põhiline geneetilist mitmekesisust säilitav evolutsiooniline jõud.
Geneetilise mitmekesisuse määratlemine
Geneetilist mitmekesisust võib defineerida ja hinnata mitmeti. Levinumad geneetilise mitmekesisuse mõõdupuud on järgmised:
- alleelide mitmekesisus – ühendab informatsiooni alleelide arvu ja suhtelise sageduse kohta lookuses. Seda mõõdetakse tavaliselt molekulaarsete markerite abil, mis on eeldatavasti neutraalsed;
- alleelide keskmine arv lookuses;
- genotüüpide arv populatsioonis;
- heterosügootsus – kaht erinevat alleeli sisaldavate lookuste protsent;
- nukleotiidide mitmekesisus – keskmine nukleotiidsete erinevuste arv lookuses kahel populatsioonist juhuslikult valitud isendil;
- polümorfsete lookuste protsent;
- geneetiline dispersioon ehk muutlikkus – muutlikkus fenotüübis avalduva tunnuse osas, mis on põhjustatud geneetilistest erinevustest;
- päritavus – kogu geneetilise muutlikkuse suhe kogu muutlikkusesse.

Uurimismeetodid
Valkude polümorfismi uurimine
1955. aastal võeti kasutusele valkude elektroforees, tehnika, mis võimaldab eraldada proteiine nende erineva liikuvuse järgi elektriväljas. Ensüümi variante, mis erinesid liikuvuse poolest elektroforeesil, aga olid kodeeritud sama lookuse poolt, hakati nimetama allosüümideks. Allosüümide kasutamisel populatsioonide geneetilise mitmekesisuse uuringutes ilmnesid aga mõningad puudused. Selgus, et tegelik geneetiline varieeruvus jääb avastamata geneetilise koodi kõdumise tõttu. Sünonüümsed nukleotiidide asendused, nn vaikivad mutatsioonid, muudavad koodoni DNA järjestust, aga ei põhjusta aminohappe asendumist ja seega ei mõjuta valku. Ka siis, kui on tegemist mittesünonüümse nukleotiidi asendusega, mis muudab aminohappelist järjestust, on uus alleel elektroforeesil tuvastatav ainult siis, kui ta on muutnud elektrilaengut, massi või struktuuri. Paljudel aminohappe asendustel sellist mõju pole.
DNA polümorfismi uurimine
Edasised arengud geneetiliste erinevuste uurimises võimaldasid otsesemalt analüüsida DNA järjestusi. Esimene selline tehnoloogia oli RFLP, mille puhul võrreldi restriktsiooni fragmentide pikkuse erinevusi. Mõjukaimaks abivahendiks geneetilise mitmekesisuse mõõtmisel sai PCR. Võrdlev genoomika, mis võimaldab hinnata erinevate taksonoomiliste gruppide sugulust geneetiliste erinevuste põhjal, on teinud suuri arengusamme. Mitmesugustel organismidel sekveneeritud geenijärjestuste kohta on koostatud mahukaid andmebaase. Allpool on toodud levinumad DNA mitmekesisuse uurimiseks kasutatavad meetodid.
- RFLP (restriction fragment length polymorphism) – restriktsioonifragmentide pikkuse polümorfism. Meetod selleks, et identifitseerida polümorfismi DNA järjestustes, kasutades restriktsiooni ensüüme ja geelelektroforeesi. Mutatsioone sisaldava DNA lõikamisel restriktaasiga tekivad teistsuguse pikkusega fragmendid võrreldes esialgse DNA-ga.
- RAPD (randomly amplified polymorphic DNA) – juhuslikult amplifitseeritud polümorfne DNA. DNA järjestustes esineva polümorfismi tuvastamiseks kasutatakse PCR-is suvalisi praimereid ja produkte uuritakse elektroforeesi abil.
- AFLP (amplified fragment length polymorphism) – amplifitseeritud fragmentide pikkuse polümorfism. Meetod põhineb DNA järjestuste polümorfismi hindamisel. Restriktsioonifragmente amplifitseeritakse PCR-i abil.
- Mini- ja mikrosatelliide uuringud. Mini- ja mikrosatelliidid on tandeemsed kordusjärjestused, mille pikkuses esineb polümorfism. Üksteisest erinevad need nukleotiidide arvu, mutatsiooniprotsesside ja kromosoomidesse jaotumise poolest, aga kahe klassi vaheline piir ei ole defineeritud.
- SNP (single mucleotide polymorphism) – erinevaid nukelotiide sisaldavate alleelide esinemine DNA järjestuse kindlas punktis.
- Mikrokiibid (microarrays) – suur hulk (tavaliselt tuhandeid) erinevaid DNA või oliginukleotiidseid järjestusi on üksteise järel ritta seatud tahkel substraadil. Kasutatakse hübridisatsiooni eksperimentides, et uurida SNP-sid ja geeniekspressiooni mustreid.
















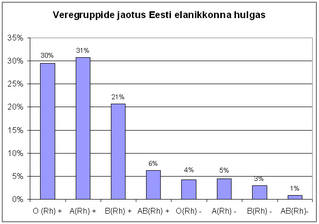






 [Public domain], via Wikimedia Commons")