Geneetika mõisted
Pärilikkus – organismide võime anda endasarnaseid järglasi.
Geneetika – teadusharu, mis uurib pärilikkust ja muutlikkust.
Geen – DNA lõik, mis määrab ühe RNA molekuli sünteesi (ja ka tavaliselt ühe tunnuse).
Alleel – geeni üks esinemisvorm (geeni erivorm). Nad asuvad homoloogiliste kromosoomide kindlas piirkonnas – lookuses.
Dominantne alleel – alleel, mille poolt määratud tunnus alati avaldub (tähistatakse suurtähega nt. A).
Retsessiivne alleel – alleel, mille poolt määratud tunnus avaldub vaid dominantse alleeli puudumisel (tähistatakse väiketähega nt. a).
Homosügoot – organism/rakk, kellel on vaadeldava tunnuse suhtes identsed alleelid, (alleelipaari seisund, kus mõlemad alleelid on kas dominantsed või retsessiivsed). Nt. AA või aa
Heterosügoot – organism/rakk, kellel on vaadeldava tunnuse suhtes erinevad alleelid (ühe geeni erinevad alleelid). Nt. Aa
Geenifond – liigi või populatsiooni geenide kogum
Genotüüp – organismi geenide kogum
Fenotüüp – organismi avaldunud tunnuste kogum
Polüalleelsus – geen esineb populatsioonis rohkem kui kahe variandina (üle kahe alleeli organismirühmas)
Polügeensus – ühte tunnust määrab palju geene
Intermediaarsus – domineerimisnähtus puudub, tunnuste vahepealne avaldumine
Kodominantsus – avalduvad mõlemad tunnused
Gregor Johann Mendel (1822-1884)
Austria munk, kes pani aluse geneetikale.
Ta tegi katseid hernestega, et selgitada tunnuste pärandumist. Valides hoolikalt, milliseid taimi omavahel ristata, avastas ta, et tunnused päranduvad paarikaupa.
Ta leidis ka, et tavaliselt avaldub fenotüübis ainult üks tunnus igast paarist.
Geneetikaülesannete sümbolid:
Mendel võttis kasutusele geneetika ülesannete ja skeemide sümbolid (kasutusel tänapäevani).
P – parentes (vanemad)
F – filia, filialis (tütar ja poeg)
x – ristamine
F1, F2, F3 – põlvkonnad.
Monohübriidne ristamine
Monohübriidse ristamise korral erinevad vanemad ühe tunnusepaari poolest.
- vaadeldakse vaid ühe tunnuse kujunemist järglastel.

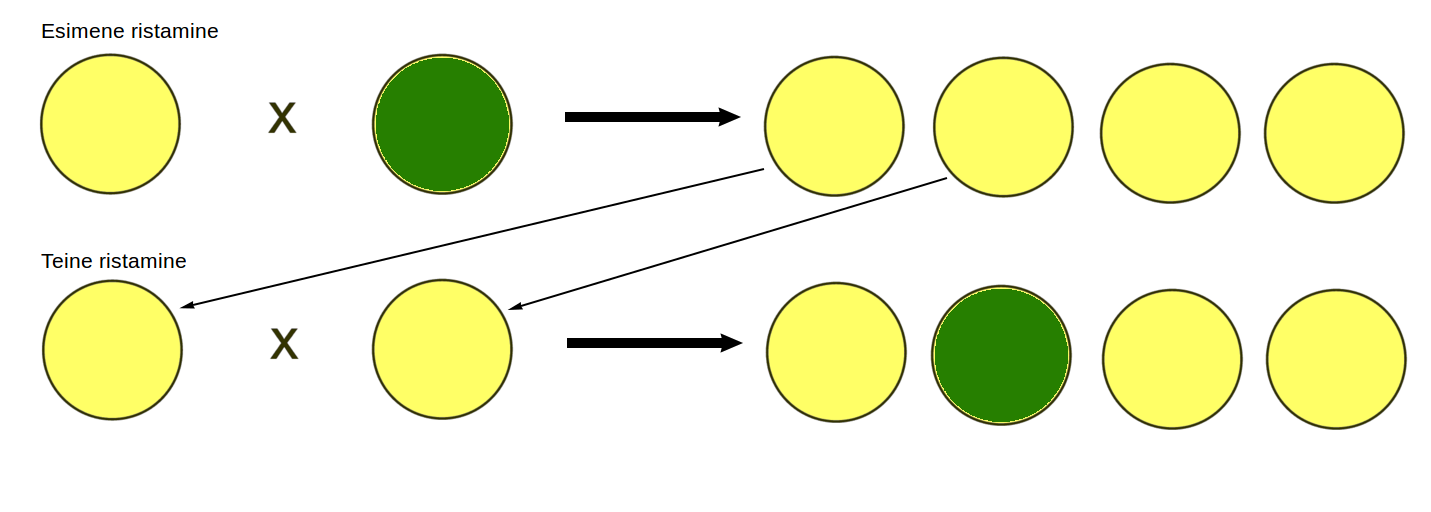
Mendeli esimene seadus ehk ühetaolisuse seadus
Homosügootide omavahelisel ristamisel moodustunud esimene järglaspõlvkond on genotüübilt samad, (heterosügootsed) ja fenotüübiliselt sarnased
sile krobeline
S-genotüüp
s-Fenotüüp
Ss-F1

siledad
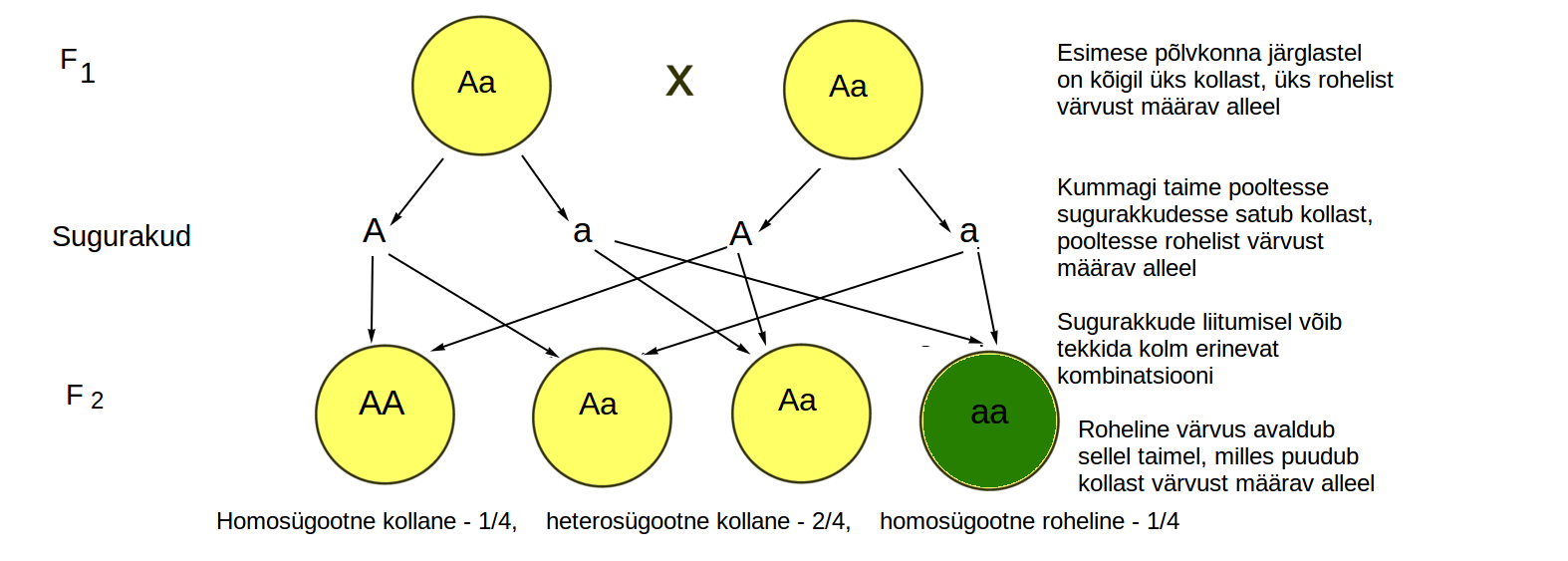
Mendeli teine ehk lahknemisseadus
Homosügootide omavahelisel ristamisel toimub järglaspõlvkonnas F2 tunnuste lahknemine seaduspäraselt (genotüübilises suhtes 1:2:1, fenotüübilt 1:2:1 või domineerimise korral 3:1 )

Järglastest genotüübilt:
1 osa on dominantseid homosügoote (BB),
2 osa heterosügoote (Bb) ja
1 osa retsessiivseid homosügoote (bb).

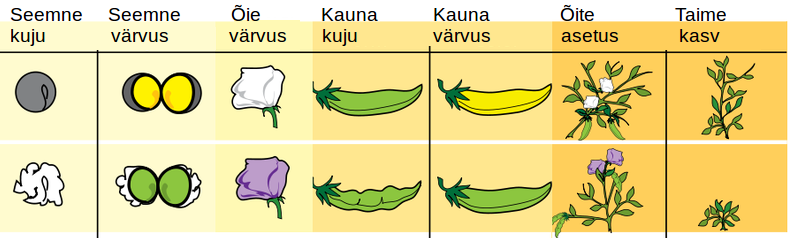
Herneste tunnused, mille pärandumise uurimise põhjal Mendel on seaduspärasused sõnastanud:

Inimese dominantsed ja retsessiivsed tunnused:

Analüüsiv meetod:
Taimede või loomade heterosügootsuse määramiseks.
Dominantse tunnusega isend ristatakse kaksikretsessiivse tunnusega isendiga.
Kui järglased on kõik dominantse tunnusega, on uuritav dominantse tunnusega isend homosügoot – sellisel juhul on tegemist tõu- või sordipuhta liigiga.
Kui järglaskonnas toimub lahknemine ja saadakse erinevate tunnustega organismid, siis järelikult on uuritav dominantse tunnusega organism selle tunnuse suhtes heterosügoot – sellisel juhul ei ole tegemist tõu- või sordipuhta liigiga.
Genealoogiline meetod:
Inimese sugupuu abil tema genotüübi väljaselgitamine.
Selgitatakse kas inimene on selle tunnuse suhtes homosügoot või heterosügoot.
Peamiselt kasutatakse pärilike haiguste riski tuvastamiseks
Inimese puhul analüüsivat ristamist kasutada ei saa.
Ülesanded:
Sinisilmne mees abiellus pruunisilmse (homosügoot) naisega. Mis värvi silmadega sünnivad nende lapsed? Koosta ristamise genotüübiline skeem!

P: aa x AA
a - sinisilmne
A - pruunisilmne
Vastus: Lapsed on kõik pruunisilmsed
2. ülesanne:
Pruunisilmne mees ja pruunisilmne naine (mõlemad on heterosügoodid) saavad lapse. Kui suur on tõenäosus, et lapsel on sinised silmad? Koosta ristamise genotüübiline skeem!
3. ülesanne:Sinisilmne mees abiellus naisega, kellel on pruunid silmad. Perekonnas on 2 tütart, üks siniste, teine pruunide silmadega. Milliseid lapsi võiksid tütred saada abielludes sinisilmsete meestega, kui pruun silmavärvus domineerib sinise üle? Koosta ristamise genotüübiline skeem.
4.ülesanne:
Ristati kahte mustakarvalist merisiga. Saadi 3 poega, kellest 2 olid mustakarvalised ja üks pruunikarvaline. Koosta ristamisskeem! Kas tegemist oli tõupuhaste merisigadega?

P: Aa x Aa
Vastus: Ei ole tõupuhtad, kuna esines ka pruune sigu
5. ülesanne:Eesmiste purihammaste puudumine pärandub dominantselt. Perekonnas, kus mõlemal vanemal esines nimetatud puue, sündis normaalne tütar. Kui suur on tõenäosus, et nende järgmisel lapsel eesmised purihambad puuduvad?
6.ülesanne:
Nudipäist veisetõugu ristati sarvilise tõuga. Hübriidid olid kõik nudid. Neid ristati omavahel ja F2 põlvkonnas saadi 52 vasikat. Milline on teoreetiline oodatav arvuline lahknemine genotüüpide ja fenotüüpide järgi?
7.ülesanne:
Paaritades nudi pulliga (dom. tunnus) sarvilisi lehmi saadi Ühes karjas 117 nudi ja 121 sarvedega vasikat. Joonistada genotüübiline skeem. Kas kahe sarvedega looma ristamisel võib saada nudisid järglasi? Nudide ristamisel sarvedega järglasi? Tehke vastavad skeemid!
Intermediaarsus:
8.ülesanne:
Ristatakse punast lillhernesorti valgeõielise sordiga. F1 põlvkonnas on kõik hübriidid roosaõielised. Milliseid lillherneid on oodata F2 põlvkonnas? Koosta ristamisskeem!
9.ülesanne: Albinismigeen domineerib valge värvuse üle. Missugune tuleb genotüübiline ja fenotüübiline lahknemine F1 põlvkonnas kui omavahel ristati kahte albiinot valge värvusgeeni suhtes heterosügootset hobust? (On teada, et homosügootsed albiinod hukkuvad juba looteeas).
10.ülesanne: Kui suur on tõenäosus, et heterosügootsete pruunisilmsete vanemate kahest lapsest on üks sinisilmne ja teine pruunisilmne?
11. Ristatakse punast lillhernesorti valgeõielise sordiga. F1 põlvkonnas on kõik hübriidid roosaõielised. Milliseid lillherneid on oodata F2 põlvkonnas? Koosta ristamisskeem!

P: ĀĀ x aa
aa - valge
ĀĀ - punane
12. ülesanne: Kodominantsus: Ristatakse valge kana ja pruun kukk. F1 põlvkonnas on kõik tibud pruuni-valgekirjud. Milliseid tibusid on oodata F2 põlvkonnas, kui F1 pruunivalge kana ristatakse pruuni kukega? Koosta ristamisskeem!
13.ülesanne: Punaseviljalise aedmaasika ristamisel valgeviljalise sordiga on F1 hübriidid roosaviljalised. Milliseid tulemusi on oodata F1 hübriidi ristamisel punaseviljalise sordiga?
14.ülesanne: Peres on vanemad laineliste juustega (Āa). Kui suur on tõenäosus, et sellesse perre sünnivad järgnevate fenotüüpidega lapsed:
a) laineliste juustega
b) lokkisjuukseline või laineliste juustega
c) sirgete juustega tütar?

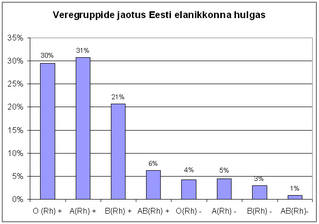






 [Public domain], via Wikimedia Commons")